Full Length Research Paper

ABSTRACT
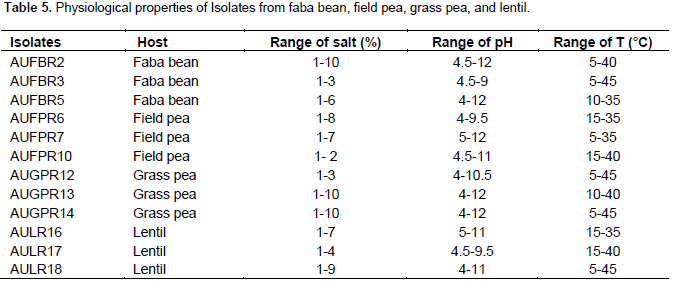
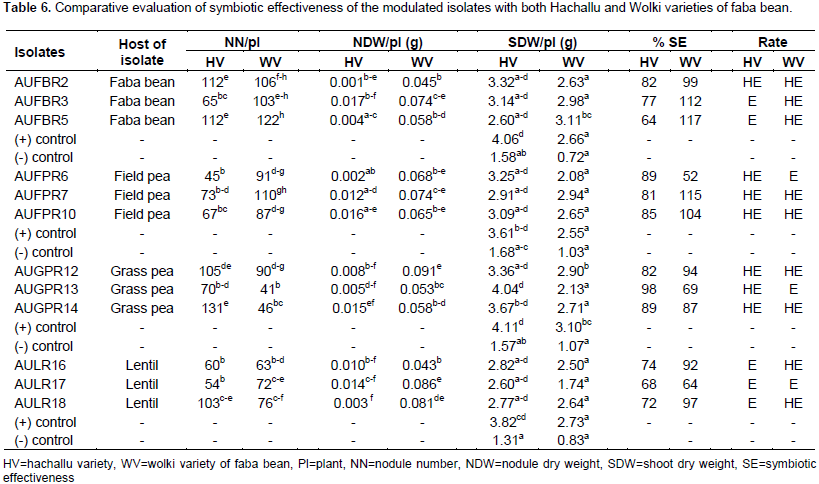
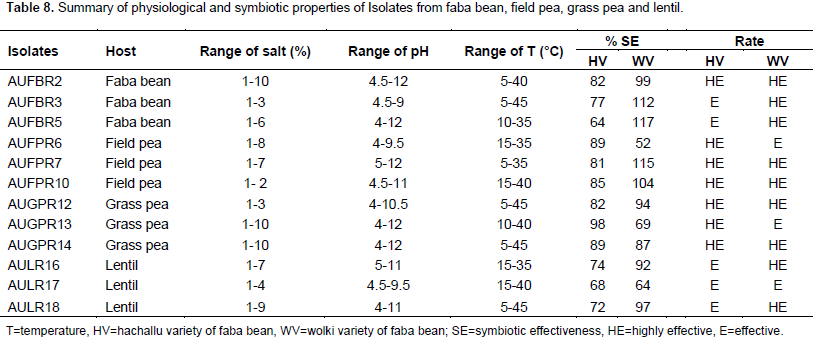
Faba bean (Vicia faba L.) is one of the most widely cultivated leguminous crops grown in the Highlands of Ethiopian. This study was aimed at evaluating the effect of Rhizobium leguminosarum bv.viciae isolates from other cross-inoculation hosts from field pea, grass pea, and lentil on faba bean varieties. Thus, 12 isolates were selected from faba bean, field pea, grass pea and lentil using host trap method on soil samples that was collected from North Shoa. All isolates were tolerated to pH (5 to 9), salt concentration (1 to 2%), and at a temperature of (15 to 35°C). Cross-inoculation experiments were conducted in the greenhouse to test the effects of R. leguminosarum isolates from faba bean, field pea, grass pea and lentil hosts on Hachallu and Wolki varieties of faba bean. The mean nodule number, nodule dry weight and mean shoot dry weight of the inoculated plants showed variations among the different cross inoculants (p<0.01). Symbiotic effectiveness varied from a minimum of 64% on Hachallu variety inoculated by isolate AUFBR5 (faba bean) and 52% on Wolki variety by isolate AUFPR1 (field pea) association to a maximum of 98% in Hachallu variety inoculated with isolate AUGPR13 (grass pea), and 117% in Wolki variety inoculated with isolate AUFBR5 (faba bean). The study showed physiological and symbiotic diversity of R. leguminosarum isolates of faba bean, field pea, grass pea and lentil. The effective isolates that have been tested with Hachallu and Wolki varities at green house level should be re-tested under field conditions to ascertain their performance in vivo.
Key words: Field pea, grass pea, lentil, symbiotic effectiveness.
INTRODUCTION
MATERIALS AND METHODS
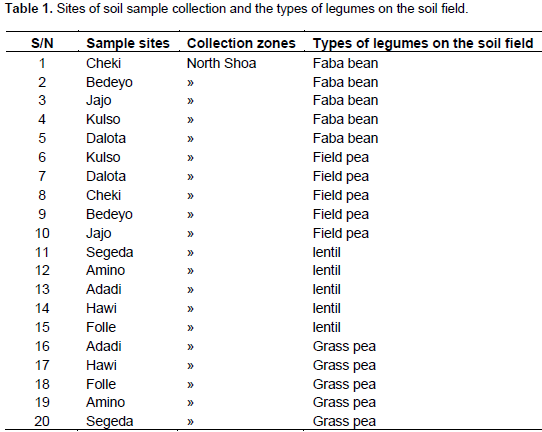
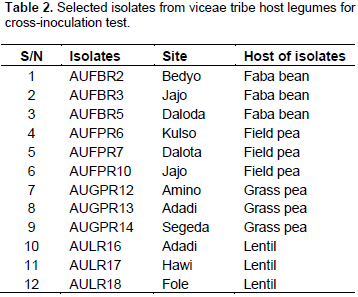

RESULTS AND DISCUSSION
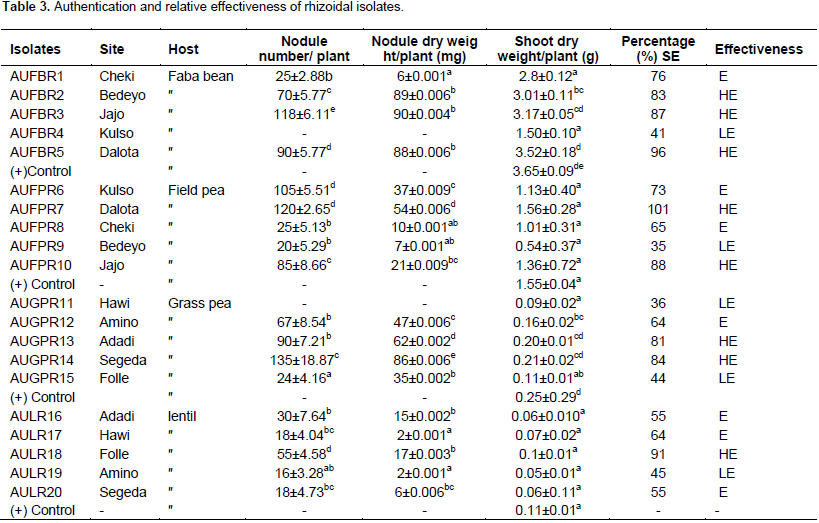
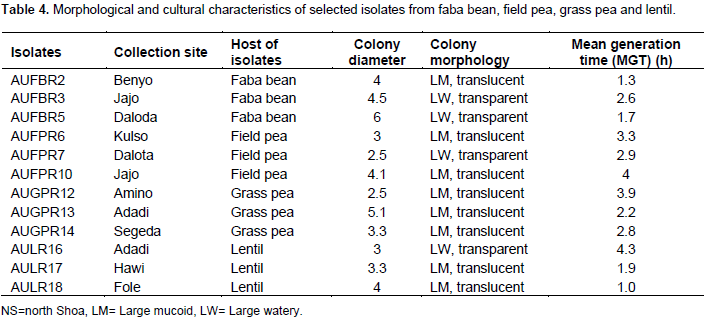

CONCLUSION
RECOMMENDATIONS
CONFLICT OF INTERESTS
ACKNOWLEDGEMENTS
REFERENCES
|
Adal M (2009). Phenotypic and Symbiotic characterization of grass pea (Lathyrus sativus) rhizobial isolates from some major growing areas of South Wollo and West Shoa, Ethiopia. MSc. Thesis. Addis Ababa University, Ethiopia. |
|
|
Adamu A, Assefa F, Hailemariam A, Bekele E (2001). Studies of Rhizobium inoculation and fertilizer treatment on growth and production of faba bean (Vicia faba) in some yield depleted and yield sustained regions of Semien Showa. SINET: Ethiop. J. Sci. 24:197-211. |
|
|
Adler K (2008). Exploring the implications of introducing inoculated legumes in southern Ethiopia: A systemic analysis of the factors affecting farmer adoption and nitrogen synchrony. M.Sc. thesis, Norwegian University of life Sciences, Norway. Available at: http://oe.confolio.org/scam/34/resource/162 |
|
|
Amara DS (1990). Evidence of cross-inoculation among the fast-growing tree legumes used in agroforestry. In: Nitrogen Fixation: Achievement and Objectives. (Gresshoff PM, Roth LE, Stacy G and Newton WE. eds). Chapman Hall, New York. P 645. |
|
|
Amsalu A (2007). Symbiotic and Phenotypic Characterization of Rhizobium leguminosarum biovar viciae isolates of Field Pea (Pisum sativum) form Different Pulse Growing Regions of Ethiopia. M.Sc. Thesis. Addis Ababa University, Addis Ababa. |
|
|
Belay Z (2006). Symbiotic and phenotypic diversity of Rhizobium leguminosarum var viceae Isolates (Vicia faba) fromNorthern Gondar, Ethiopia. Msc. Thesis. AA.University, pp. 7-8. |
|
|
Belay Z, Assefa F (2011). Symbiotic and phenotypic diversity of Rhizobium leguminosarum bv.Viciae from Northern Gondar, Ethiopia. Afr. J. Biotechnol. 10(21):4372-4379. |
|
|
Bernal G, Graham PH (2001). Diversity in the rhizobia associated with Phaseolus vulgaris L. in Ecuador, and comparisons with Mexican bean rhizobia. Can. J. Microbiol. 47:526-534. |
|
|
Brockwell J (1998). Matching rhizobia and temperate species of Acacia. In: Recent Development in Acacia planting. (Turn bull, LW, Crompton, HR and Pinyopusarerk K. eds). ACIAR, Canberra, Australia pp. 264-273. |
|
|
Central Statistics Authority (CSA) (2000). Agricultural Sample Survey. 2000/2001. Report on area and production for major crops. Central Statistical Authority, Addis Ababa, Ethiopia. Stat. Bull. P 245. |
|
|
Central Statistics Authority (CSA) (2005). Agricultural Sampling Survey 2004/2005. Report On Area and Production for Major Crops, Addis Ababa, Ethiopia pp. 22-34. |
|
|
Central Statistics Authority (CSA) (2015). The federal democratic republic of Ethiopia central statistical agency agricultural sample survey 2014/2015. Area and production of major crops (Private peasant holdings, Meher season). Addis Ababa, Ethiopia: Central Statistical Agency P 19. |
|
|
Demissie N (2006). Genetic diversity and characterization of indigenous Rhizobium leguminosarum biovar vicia isolates of cool-season food Legumes grown in the highlands of Ethiopia. Ph.D. Thesis, University of Putra, Malasia pp. 26-35. |
|
|
Fano B (2010). Phenotypic and Symbiotic characteristics of Rhizobia nodulating field Pea (Pisum sativum L.) in southern Tigray, Ethiopia, M.Sc. Thesis. Addis Ababa University, Addis Ababa. |
|
|
Gebremariam G (2007). Characterization and evaluation of nitrogen fixation of Rhizobium leguminosarum bv. Viciae isolates collected from different parts of Ethiopia. MSc. Thesis. Addis Ababa University, Ethiopia. |
|
|
Giller KE (2001). Nitrogen fixation in tropical cropping systems 2nd ed. CABI, international, Wallingford. |
|
|
Hailemariam A, Tsigie A (2006). Symbiotic nitrogen fixation research on food legumes in Ethiopia. In: Proceedings of a Workshop on Food and Forage Legumes (Kemal A, Gemechu K, Seid A, Rajendra M, Surendra B, and Khaled M eds). ICARDA, Addis Ababa, Ethiopia. pp. 172-176. |
|
|
Hailemariam A, Tsigie A (2006). Symbiotic nitrogen fixation research on food legumes in Ethiopia. In: Proceedings of a Workshop on Food and Forage Legumes (Kemal A, Gemechu K, Seid A, Rajendra M, Surendra B, and Khaled M eds). ICARDA, Addis Ababa, Ethiopia. pp. 172-176. |
|
|
Handley BA, Hedges AJ, Beringer JE (1998). Importance of host plants for detecting the population diversity of Rhizobium leguminosarum biovar viciae in soil. Soil Biol. Biochem. 30:241-249. |
|
|
Ihsanullah D, Hasan S, Khan BM, Gul H, Ijaz AKh (2008). Effect of Different Levels of Nitrogen on Dry Matter and Grain Yield of Faba Bean (Vicia faba L.). Pak. J. Bot. 40(6):2453-2459. |
|
|
Jensen ES, Peoples M, Hauggaard-Nielsen H (2010). Review: faba beans in cropping systems. Field Crop Res.115:203-216. |
|
|
Jordan D (1984). Rhizobaceae. In: Bergey's Manual of Systematic Bacteriology. (Hendricks DP, Sneath HA and Halt JH, eds). Orient Longman, New York pp. 234-256. |
|
|
Keneni A, Assefa F, Prabu P (2010). Characterization of acid and salt tolerant Rhizobial strains isolated from Faba bean Fields of Wollo, Northern Ethiopia. J. Agric. Sci. Technol.12:365-376. |
|
|
Laguerre G, Louvrier P, Allard MR, Amarger N (2003). Compatibility of Rhizobial Genotypes within Natural Populations of Rhizobium leguminosarum Biovar viciae for Nodulation of Host Legumes. Appl. Environ. Microbiol. 69(4):2276-2283. |
|
|
Lo´ pez-Bellido L, Lo´ pez-Bellido RJ, Redondo R, Benı´tez J (2006). Faba bean nitrogen fixation in a wheat-based rotation under rainfed Mediterranean conditions: effect of tillage system. Field Crops Res. 98:253-260. |
|
|
Lupwayi N, Haque I (1994). Legume-Rhizobium Technology Manual. Environmental Sciences Division International Livestock Center for Africa. Addis Ababa, Ethiopia pp. 1-93. |
|
|
Mahajan A, Gupta RD (2009). Intrernational Nutrient Management in Sustainable Rice Wheat Cropping System. Springer Science and Business Media B.V., India pp. 79-80. |
|
|
Martinez-Romero E, Segovia L, Mercante FM, Franco AA, Graham P, Pardo MA (1991). Rhizobium tropici, a novel species nodulating Phaseolus vulgaris L. beans and Leucaena sp trees. Int. J. Syst. Bacteriol. 41:417-426. |
|
|
McVicar R, Panchuk K, Brenzil C, Hartley S, Pearse P (2005). Faba bean in Saskatchewan. Saskatchewan Agriculture, Food, and Rural Revitalization. University of Saskatchewan, Vandenberg P 11. |
|
|
Mulongoy K (2004). Technical paper 2: Biological nitrogen fixation. Can. J. Microbiol. 21:1-19. |
|
|
Nikfarjam SG, Aminpanah H (2015). Effects of phosphorus fertilization and Pseudomonas fluorescence strain on the growth and yield of faba bean (Vicia faba L.). IDESIA 33(4):5-21. |
|
|
People MD (1995). Biological nitrogen ï¬xation: An efficient source of nitrogen for Sustainable Agricultural production. J. Soil Sci. 174:3-8. |
|
|
Purcino HMA, Festin PM, Elkan GH (2000). Identification of effective strains of Brad rhizobium for Archis Pintoi. Trop. Agric. 77:226-231. |
|
|
Salunkhe DK, Kadam SS (1989). Handbook of world food legumes. In: Nutrition chemistry, processing technology, and utilization. Boca Rotan, Florida: CRC press, Inc. P. 310. |
|
|
Shimekit F, Mengistu A, and Assefa F (2008). Pattern of Nodulation and Nitrogen Fixing Performance of Some Introduced Forage Legumes of Ethiopia. Ethiop. J. Nat. Resour. 10:161-176. |
|
|
Siddique KH, Johansen C, Turner NC, Jeuffroy MH, Hashem A, Sakar D, Gan Y, Alghamdi SS (2012). Innovations in agronomy for food legumes: a review. Agron. Sustain. Dev. 32:45-64. |
|
|
Somasegaren P, Hoben HJ (1994). Handbook for Rhizobia: Methods in legume rhizobium technology. Springer-Verlay, New York. |
|
|
Talukder MS, Solaiman A, Khanam D, Rabbani MG (2008). Characterization of Some Rhizobium Isolates and Their Effectiveness on Pea. Bangl. J. Microbiol. 25:45-48. |
|
|
Tesfaye G (2008). Symbiotic and phenotypic diversity of rhizobial isolates nodulating Vicia faba from western Shoa and Hararghe, Ethiopia. M.SC. Thesis. Addis Ababa University, Ethiopia. |
|
|
Teshome Z (2006). Numerical taxonomy of phenotypic characters of Rhizobium leguminosarum var vicieae isolates from the major pulse producing regions of Ethiopia and its importance for predictive ecological and symbiotic studies. MSc. Thesis. Addis Ababa University, Addis Ababa. |
|
|
Tran LSP, Nguyen HT (2009). Future Biotechnology of Legumes. In: Emerich WD, and Krishnan H, eds., Nitrogen Fixation in Crop Production, The American Society of Agronomy, Crop Science Society of America and Soil Science Society of America, Madison pp. 265-308. |
|
|
Vincent JM (1970). A Manual for the Practical Study of Root Nodule Bacteria. Blackwell Scientific Publications, Oxford P 164. |
|
|
White D (1995). The physiology and biochemistry of prokaryotes. Oxford University Press, Oxford. |
|
|
Yohannes D (2000). Faba bean (Vicia faba) in Ethiopia. Institute of biodiversity, conservation and Research (IBCR), Addis Ababa, Ethiopia P 43. |
|
|
Zhange X, Kosier B, and Priefer UB (2001). Symbiotic plasmid rearrangement in Rhizobium leguminosarum bv. Viciae VF39SM. J. Bacteriol. 183: 2141-2144. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


