Full Length Research Paper

ABSTRACT
This study aims to characterize chemical groups of metabolites in extracts of Acanthospermum hispidum and Croton zambesicus, and to evaluate their anti-inflammatory activities and their immunomodulatory effect on two pro-inflammatory cytokines (TNFα and IL6). The phytochemical screening was done on the powders of both plants by staining and precipitation reactions. Anti-inflammatory activity was performed in vivo on rat paw edema induced by 2% formalin and immunomodulatory activity was performed in vitro on rat Peripheral Blood Mononuclear Cells (PBMC) stimulated by Lipopolysaccharide (LPS). The leaves of the plants contained important chemical groups such as: cachectic tannins, flavonoids, terpenes, saponosides and quinones. The ethanolic extracts of the two plants studied showed good anti-inflammatory activity by reducing the volume of the rat paw edema induced by 2% formalin and by inhibiting the production of TNFα, IL6 by PBMC of rats stimulated by LPS.
Key words: Acanthospermum hispidum, Croton zambesicus, extracts, phytochemical screening, biological activities.
INTRODUCTION
The healing properties of certain medicinal plants have been researched, noted, and passed down from generation to generation (Yapi, 2015). The continuous and perpetual human interest in medicinal plants has led to the modern and sophisticated treatment of several human diseases (Petrovska, 2012). The World Health Organization (WHO) has estimated that 80% of the world's population uses herbal formulations as traditional therapies (WHO, 2002). Faced with the therapeutic limitations of modern chemically formulated drugs, the development of herbal research has shifted to the formulation of phytomedicines or traditionally improved the development of herbal research has shifted to the formulation of phytomedicines or traditionally improved drugs to better treat human pathologies (Guedje et al., 2012).
Thus, herbal medicine is a good alternative to conventional drugs as it has been used for centuries to treat human diseases (Ambe et al., 2015). It is mainly based on the idea that plants are a natural means of treatment devoid of any risks. The consumers often believe that natural is synonymous with harmless. However, a plant can be both useful and toxic. Some compounds in the secondary metabolism of plants are serious poisons, such as cyanogen derivatives (Agbankpe et al., 2015). Several studies have highlighted the pharmacological effects of many plants. However, it is also important to evaluate the toxicity of herbal preparations to determine the safety of these remedies (Ouahchia et al., 2017). An ethnobotanical study of plants used in the treatment of inflammation among herbalists in the market in southern Benin has identified a number of medicinal plants with anti-inflammatory properties (Kpodji et al., 2019). In this study, Acanthospermum hispidum and Croton zambesicus were among the plants with a high citation frequency that were selected.
A. hispidum belongs to the Asteraceae family and the study of the literature reveals those various parts of A. hispidium have been used as a folklore medicine to treat various diseases (Roy et al., 2010; Chakraborty et al., 2012).
C. zambesicus belongs to the genus Euphorbiaceae, which includes about 1,300 species. It is widespread in tropical regions, both in the old and new world. Several species have been used for a long time as medicinal plants in Africa, Asia and South America. Popular uses include the treatment of several diseases such as cancer, digestive problems, and inflammation (Salatino et al., 2007). In Benin, the decoction of the leaves of C. zambesicus is used as an antihypertensive and in treatment of malaria-related fever (Block et al., 2004).
Most research on medicinal plants focuses on the characterization of secondary metabolites that have shown significant biological activities. Secondary metabolites represent a large group of biologically active compounds that are synthesised as a defense mechanism of plants against various microorganisms, insects, herbivores and different environmental extremes, as well as for protection against UV light (Xie and Lou, 2009; Klavina, 2018). Secondary metabolites such as phenolics, flavonoids and terpenoids, have been shown to have antioxidant, anti-inflammatory, antitumour and antibacterial effects (Klavina et al., 2015; Lunic et al., 2020; Mandic et al., 2021).
The aim of this study was to investigate the potential bioactive effects of ethanolic extracts of A. hispidum and C. zambesicus, under in vitro conditions. By knowing these processes, we obtain new knowledge that can be applied to treat certain pathological conditions. Thus, this study proposes to perform preliminary phytochemistry tests to detect secondary metabolisms (unpurified compounds) present in ethanolic extracts of A.hispidum and C. zambesicus and to evaluate their anti-inflammatory properties and their immunomodulatory effect on pro-inflammatory cytokines TNFα and IL6.
MATERIALS AND METHODS
Plant material
The plant material was the leafy stems of A. hispidum and C. zambesicus. The plants were harvested in Lokossa (Latitude: 6.63333, Longitude: 1.71667 6° 37′ 60″ N, 1° 43′ 0″ E) in the Department of Mono in south-western Benin in January 2019. The two species of plants selected were identified with the national herbarium of the University of Abomey-Calavi following the numbers YH516/HBN for A. hispidium and YH517/HBN for C. zambesicus. The leafy stems were dried in the laboratory at room temperature in a covered space. After drying, the plants were ground and powdered at the mill. The powders were stored in sealed glass jars for further analysis.
Biological material
To assess larval toxicity Artemia salina shrimp larvae were used. A. salina’ eggs (ARTEMIO JBL D-67141 Gmbh Neuhofem), acquired from the Applied Hydrobiology’s Laboratory of Agricultural Sciences’ Faculty at University of Abomey-Calavi (UAC, Benin). Rats' PBMCs were used to assess immunomodulatory activity. Male Wistar rats aged at least three months with a body weight between 163 g and 209 g were used. These animals were acquired at the Animal Farm of the Institute of Applied Biomedical Sciences of the University of Abomey-Calavi of Benin. They were housed in cages covered with wood chips and acclimatized for 2 weeks before experiment at the animal farm of the Research Unit in Applied Microbiology and Pharmacology of Natural Substances (URMAPha). The rats had free access to water and food. Animal Research Review Panel and Animal Welfare Unit regulations of temperature and lighting systems were maintained with a room temperature of 20–26°C and regular light cycles of 12 h light/dark. All methods and protocols used in this study were observed following established public health guidelines “Guide for Care and Use of Laboratory Animals”.
Ethical approval
This study is part of a thesis. The Committee of the Doctoral School of Life and Earth Sciences (ED-SVT) of the University of Abomey-Calavi (UAC-Benin) under the number 10185509 has authorized this study on October 10, 2018 for a period of 3 years.
Ethanolic extraction of two plants
The two plant species were extracted with ethanol, a method that is an adaptation of the protocol used by Okokon et al. (2010). It has the advantage of putting the powder properly in contact with the solvent through continuous agitation. A 50 g mass of leafy stems powder was macerated in 500 mL of ethanol 96% under continuous agitation for 72 h. The mixture was filtered three times on hydrophilic cotton and once on Whatman No1 filter paper. The filtrate was evaporated at a temperature of 40°C in an oven until a dry mass of the ethanol extract was obtained. The yield of the raw extract is defined as the ratio between the mass of the dry extract and the mass of plant powder treated (Deguenon et al., 2017). This return (R) was calculated by the formula:
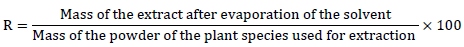
The extracts were stored at 4°C in closed glass jars until used.
Phytochemical screening
Coloring and precipitation reactions were used to characterize the chemical groups present in the extract. The methods cited by Bruneton (1999) were used as described by Houngbeme et al. (2014) and Okokon and Nwafor (2010).
Larval cytotoxicity test
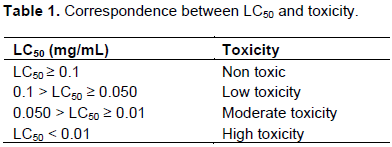
The cytotoxic effect of the ethanolic extracts was evaluated on brine shrimp larvae according to the method described by Dougnon et al. (2013). A. salina larvae were obtained by hatching 10 mg of A. salina eggs under continuous agitation in 1 L of seawater for 48 h. A 20 mg/mL concentrated stock solution was prepared for each plant extract. A series of dilutions of each stock solution was carried out to obtain a decreasing concentration. 1 mL of each diluted solutions was added to 1 mL of sea water containing 16 live larvae. A control solution without the extract was prepared under the same conditions. All solutions were incubated under agitation for 24 h at ambient temperature. Dead larvae were counted using an optical microscope to get a representative curve of the number of surviving larvae versus the concentration of the extract. The data (concentration-response) were log transformed, and the mean lethal concentration (LC50) was determined. To assess the larval toxicity of the extract, the correlation grid associating the degree of toxicity with LC50 proposed by Mousseux (1995) was used (Table 1).
In vivo anti-inflammatory activity test
Anti-inflammatory activity by reduction in the volume of edema was carried out by the adapted method of Winter et al. (1962). Inflammation is induced by injection of carragenan at the plantar arch of the rat's right leg. In this protocol, the phlogogenic agent used is formalin at 2%. The edema caused by this phlogogenic agent is translated into volume and measured by the "digitally displayed sliding foot"; which allows us to follow the evolution of the inflammatory process. Five lots of six rats were used: - Lot 1 (negative control): Rats in this lot received no treatment. The values measured here are natural physiological values; - Lot 2 (positive control): rats in this lot received only 2% formalin (100 µl; 2%) in the arch of the rat's right leg; - Lot 3 (Reference Lot): Rats in this lot were treated orally with diclofenac at a rate of 50 mg/kg, 1 h before formalin injection; - Lot 4: Rats in this lot received the ethanol extract of C. zambesicus orally, at a rate of 200 mg/kg, 1 h before the injection of formalin; - Lot 5: Rats in this lot received the ethanol extract of A. hispidum orally, at a rate of 200 mg/kg, 1 h before the injection of formalin. The evolution of edema was tracked by measuring the legs. Leg volume was measured at T0 (before formalin injection), T1 (1 h), T2 (2 h), T3 (3 h), T4 (4 h) and T5 (5 h) after the formalin injection. The change in leg volume was calculated by making the difference between the value measured at Tx (measurement time) and T0.
Immunomodulatory activity of ethanol extracts of A. hispidum and C. zambesicus
The rats were randomly distributed in three groups of three rats. In group 1, rats received the same volume of water per oral feeding as those given to animals treated with extracts. In groups 2 and 3, rats received either A. hispidum ethanol extract or C. zambesicus at a dose of 200 mg/kg of body weight. After 4 h, 5 ml of blood was collected from rats by puncture of the eye vein in tubes containing ethylene diamine tetra acetic acid (EDTA) as an anticoagulant.
After blood collection, mononucleated cells were separated from other blood components by centrifugation (Amro et al., 2013). Briefly, the 5 ml of blood was diluted to 1/2 with Roswell Park Memorial Institute (RPMI) medium (1640 without L-Glutamine, Sigma cat: R0883) in 50 ml sterile Falcon tubes. The RPMI was placed at room temperature (approximately 25°C) 30 min before use. After adding the RPMI, the tube is closed and the contents mixed gently by inversion 5 times. Five ml ficoll (Ref: 1066-5745, Fischer Scientific) were deposited in a new identified tube. The 10 ml of diluted blood was delicately deposited over the Ficoll solution. After a centrifugation at 800g for 30 min and at 20°C, with low acceleration and without brakes, the Peripheral Blood Mononucleated Cells (PBMCs) were recovered and washed with RPMI. After washing, viability and number of cells are determined by making a volume dilution for volume of cell suspension with trypan blue. PBMCs are then stored at a concentration of 2x106 cells/ml in an ice bath.
Stimulation and culture of rat PBMCs
PBMCs were cultivated in plates of 24 wells at a density of 2.106 cells/ml in RPMI in a wet-atmosphere oven at 37°C and 5% CO2. PBMCs of rats not fed by extracts were pre-incubated with ethanol extracts from each plant at a concentration of 50 or 100 μg/ml for 30 min. Subsequently, all PBMCs of rats in the different batches were stimulated with LPS at a concentration of 10 µg/ml and cultured supernatant were collected 4 h, 16 h and 24 h after stimulation. The supernatants were taken, centrifuged to remove cellular debris and stored at -80°C until the TNF-α and IL-6 cytokines were dosed. Indeed, LPS is a molecular pattern associated with pathogens (PAMP-Pathogen Associated Molecular Pattern). Once recognized by the TLR-4/CD14 complex (Park and Lee, 2013), it induces the secretion of pro-inflammatory cytokines by monocytes and macrophages (Amoroso et al., 2012). This ability justified the use of LPS in our study. A limitation of our methodology is that we did not test the effect of the ethanolic extract of the plants on TNF-α production by PBMCs without stimulation by LPS.
Quantification of TNF-α and IL-6
The production of TNF- α and IL-6 in the surpernatants of the cultivation was measured by ELISA with a commercial BioLegend kit: TNF-α with the LEGEND MAX™ Rat TNF-α ELISA Kit (No.438207, Biolegend, Amsterdam) and the IL-6 with the LEGEND MAX™ Rat IL-6 ELISA Kit (No.437107, Biolegend, Amsterdam). The dosage sensitivity is 4.2 pg/ml for TNF-α and 5.3 pg/ml for IL-6.
Statistical analysis
The susceptibility tests of bacterial strains were done with three replicates (n = 3) and the results were subjected to two-way ANOVA according to Turkey’s multiple comparison test, p < 0.05 Comparisons of average TNF-α and IL-6 concentrations were made with the Turkey test. The data presented are representative of three different experiments. The graphics were made with Graph Pad Prism 7 software.
RESULTS
Phytochemical screening
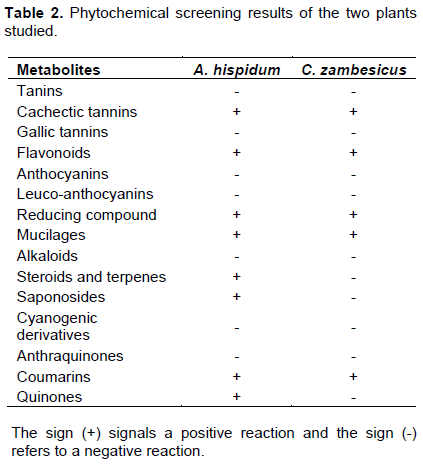
Of the 50 g of leafy stem powder of the two plants used, the ethanolic extraction of the leafy stems of A. hispidum gave a yield of 7.54% (3.77 g) and that of C. zambesicus 7.84% (3.92 g). The phytochemical analysis of the components of these two plants studied revealed the presence of chemical components in the ethanol extracts of these plants, including cachectic tannins, flavonoids, reductive compounds and others. A presence of saponosides, steroids and terpenes were found in the ethanol extract of A. hispidum. However, there is a total absence of gallic tanins, anthocyanins, leuco-anthocyanins, alkaloids, cyanogenic derivatives and anthraquinones in the ethanolic extracts of the two plants studied. The data are presented in Table 2.
Larval cytotoxicity
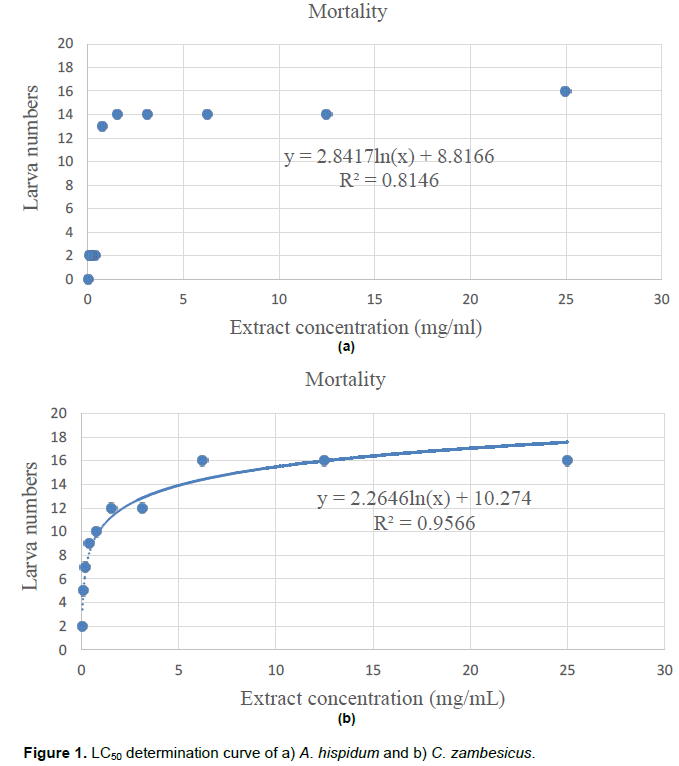
The study of A. hispidum cytotoxicity showed a lethal 50 concentration (LC50) of 0.75 mg/mL (Figure 1a) and C. zambesicus a LC50 of 0.36 mg/mL (Figure 1b). Therefore, the two extracts show no cytotoxicity.
Anti-inflammatory activity
Injection of 2% formalin caused acute edema in all rats. The percentages of increase in edema are shown in Figure 2. In control rats that were treated with physiological water, the percentages of increase in edema remained high from the first hour to the fifth hour with a maximum of 65.17 ± 0.79% at the third hour. In rats treated with extracts and the reference molecule (Diclofenac), there is a gradual decrease in percentages of increase from the first hour to the fifth hour.
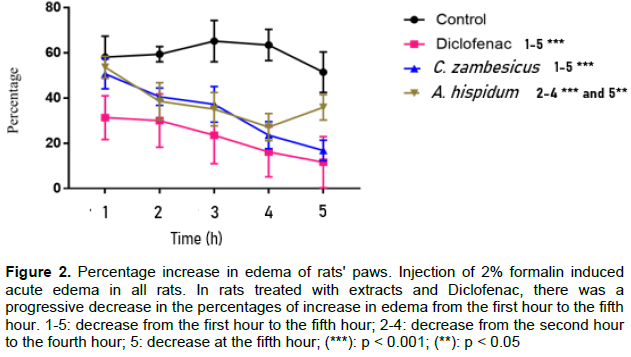
This decline was very significant (p < 0.001) in rats treated with diclofenac and C. zambesicus extract, respectively from the first and second hour. In rats treated with A. hispidium extract the decrease was very significant (p < 0.001) from the second hour to the fourth hour and significant (p < 0.05) at the fifth hour. But it should be noted that there is an increase from the fourth hour to the fifth hour. In addition, there is no significant difference between the percentages of increase in edema in rats treated with diclofenac and the two types of plant extracts used from the second hour to the fourth hour. At the fifth hour, there was a very significant difference between the percentages of increase in edema in rats treated with diclofenac and A. hispidium extract.
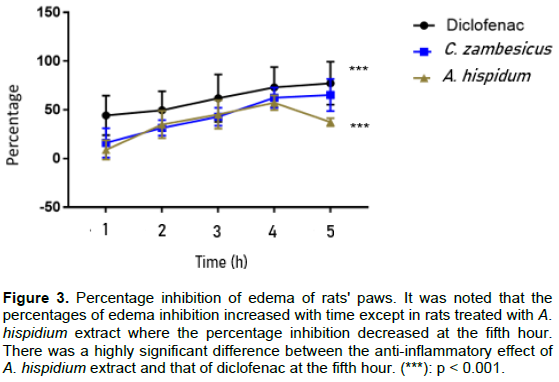
In Figure 3, the inhibition percentages of edema in rats treated with the reference molecule (Diclofenac) and the two types of plant extracts used were shown. It was noticed that the percentages of edema inhibition increased over time except in rats treated with A. hispidium extract (Ah) where the inhibition percentage decreased at the fifth hour. The inhibition percentages of edema ranged from 44.30 ± 2.99% to 77.28 ± 0.82% for diclofenac, from 16.01 ± 1.94% to 65.21 ± 8.15% for the C. zambesicus extract (Cz) and from 8.88 ± 1.53% to 57.25 ± 6.23% for A. hispidium extract between the first and fifth hour. There is no significant difference (p > 0.05) between the anti-inflammatory effect of the reference molecule used and that of the C. zambesicus extract from the second hour to the fifth hour. Also, the anti- inflammatory effect of A. hispidium extract is similar to that of diclofenac from the second hour to the fourth hour (p > 0.05), but there is a very significant difference (p < 0.001) between the anti-inflammatory effect of this extract and diclofenac at the fifth hour.
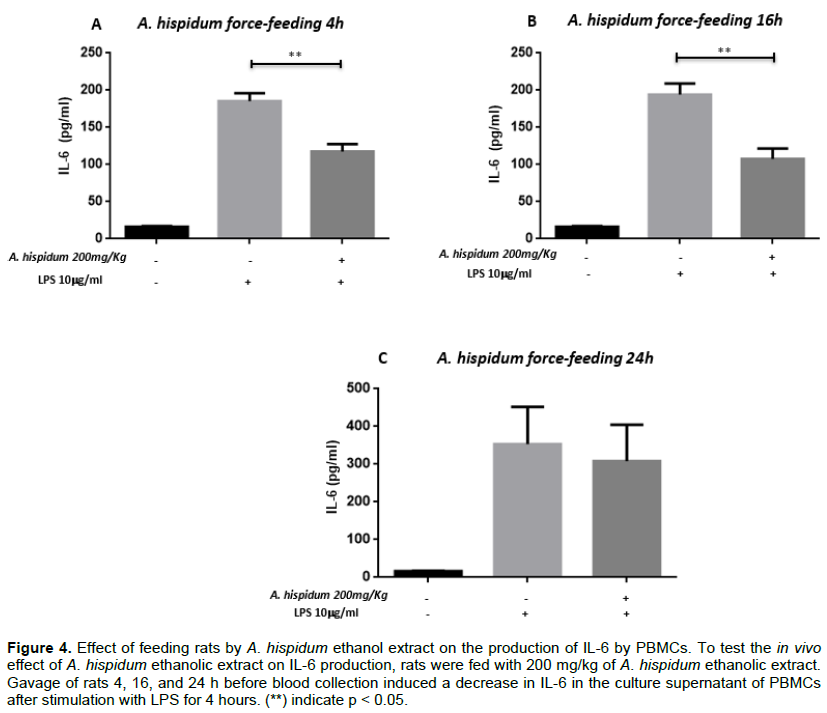
Effect of feeding rats by A. hispidum ethanol extract on the production of IL-6 by PBMCs
In order to test the in vivo effect of A. hispidum ethanol extract on IL-6 production, rats were fed with 200 mg/kg of A. hispidum ethanol extract. A gavage feeding of rats 4h before blood sampling induced a decrease in IL-6 in PBMC’s culture supernatant after stimulation with LPS for 4 h. Indeed, in the absence of gavage, the average concentration of IL-6 in the culture supernatant of mononucleated blood cells of the peripheral blood of rats was 185.453 ± 34.422 pg/ml. After gavage it was 111.820 ± 31.846 pg/ml (p = 0.014).
Similarly, PBMCs stimulated with LPS for 16 h after the gavage feeding of rats for 4 h induced a decrease in IL-6 production in PBMCs cultured supernatants. The average IL-6 concentration was 194.197 ± 26.002 pg/ml in the supernatant of PBMCs cultures of non-fed rats while it was 107.630 ± 26.002 pg/ml in that of PBMCs from fed rats (p = 0.014).
The same observations were made after 24 h stimulation of PBMCs of unfed vs. fed rats. In the supernatant of PBMCs from unfed rats, the average concentration of IL-6 was 367.500 ± 31.846 pg/ml, while it was 265.875 ± 31.846 pg/ml in fed rat PBMCs (Figure 4). This difference was not significant.
Effect of the addition of A. hispidum and C. zambesicus extracts on the production of TNF-α by LPS-stimulated PBMCs
To assess the effect of A. hispidum ethanol extract by dose and time on TNF-α production, peripheral blood mononucleated cells were pre-processed with A. hispidum ethanol extract before being stimulated with LPS at a concentration of 10 μg/ml. No significant inhibitory effects of A. hispidum extract on TNF-α production by these cells were observed at any dose and time (Figure 5). PBMCs were or were not treated with 50 μg/ml or 100 μg/ml of ethanolic extract from A. hispidum and for 30 min and stimulated with 10 μg/ml of LPS. The cultivations were collected after 4 h (A), 16 h (B) and 24 h (C). The rat TNF-α was dosed in these supernatants with the commercial TNF-α dosing kit (Figure 5).
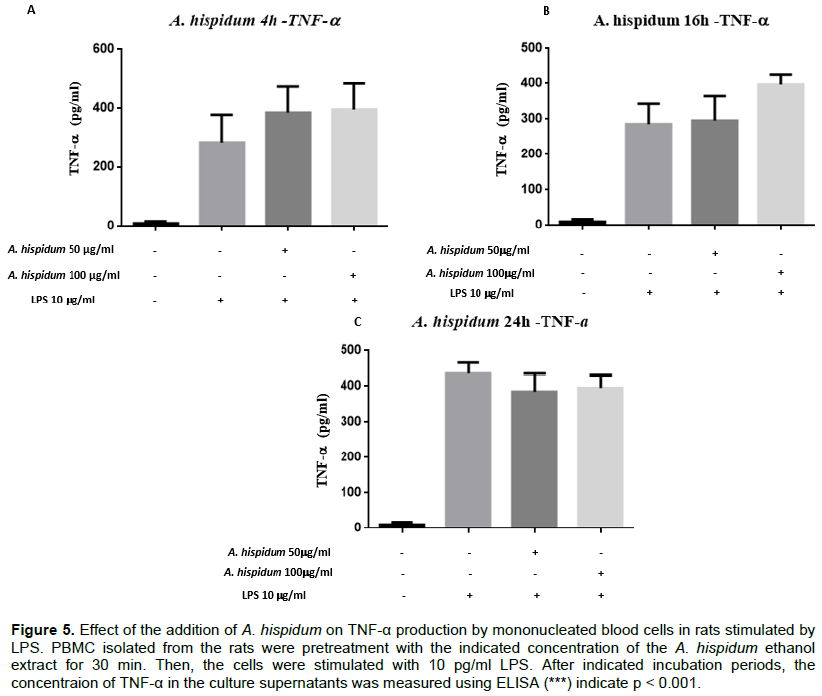
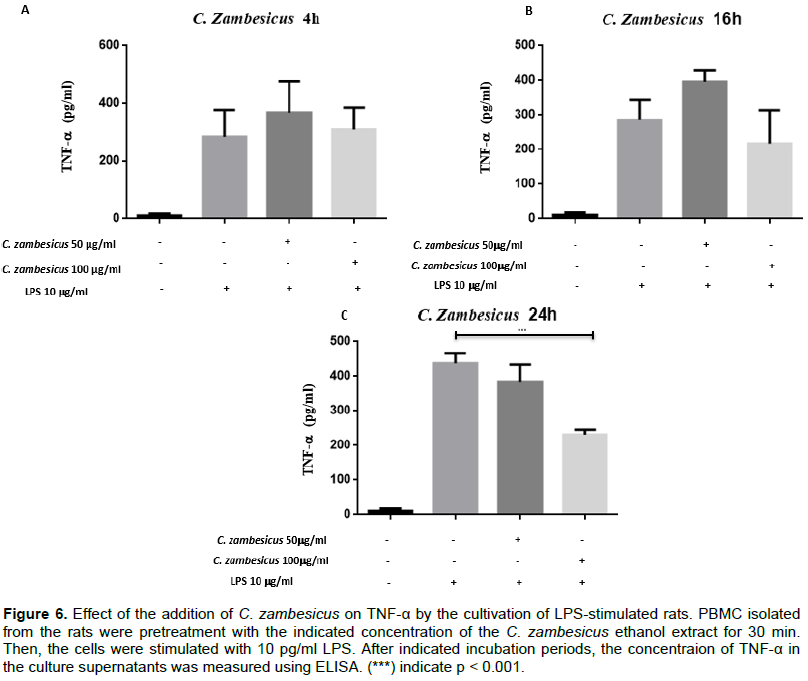
Similarly, the ethanol extract of C. zambesicus at the dosage of 50 μg/ml did not induce an inhibition of TNF-α production regardless of the time.
On the other hand, this 100 μg/ml extract resulted in a decrease in the amount of TNF-α secreted at 24 h compared to the amount of TNF-α produced by cells that were not pre-attacked. The average amount of TNF-α produced by untreated cells is 446.790 ± 41.591 pg/ml. After pre-treatment with 100 μg/ml of the extract, we observed a decrease in TNF-α production by PBMCs with an average concentration of 231.303 ± 41.591 pg/ml (p < 0.001) (Figure 6). PBMCs were or were not treated with 50 μg/ml or 100 μg/ml of C. zambesicus ethanol extract for 30 min and stimulated with 10 μg/ml of LPS. The cultivations were collected after 4 h (A), 16 h (B) and 24 h (C). Production levels in TNF-α in the supernatants of the cultivation were measured with a commercial BioLegend kit (p < 0.001) (Figure 6).
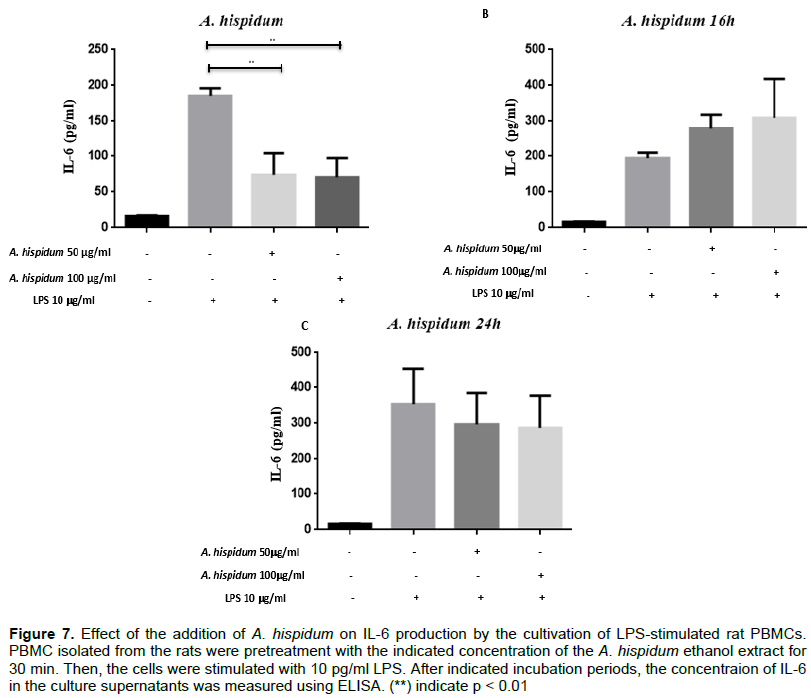
To assess the effect of ethanol extracts of A. hispidum and C. zambesicus based on dose and time on IL-6 production, PBMCs were pre-processed with these extracts for 30 min before being stimulated with LPS at a 10 μg/ml concentration. The ethanol extract of A. hispidum (Ah) at 50 μg/ml and 100 μg/ml resulted in a decrease of the quantity of IL-6 secreted at 4 h compared to the quantity of IL-6 produced by cells that were not pretreated.
The average quantity of IL-6 produced by untreated cells is 185.453 ± 34.422 pg/ml. After pre-handling with 50 and 100 μg/ml, we observed a decrease in IL-6 production by PBMCs with average concentrations of 73.780 ± 34.422 pg/ml (p = 0.002) and 83.820 ± 42.158 pg/ml (p < 0.010) respectively (Figure 7A). At the sixteenth hour, this decrease in IL-6 production by pre-treated PBMCs with the ethanol extract of A. hispidum is no longer observed. Indeed, the average quantity of IL-6 produced by untreated cells is 194.197 ± 34.422 pg/ml. The average quantities produced by pretreated cells with 50 and 100 μg/ml ethanolic extract of Ah are 277.900 ± 34.422 pg/ml and 309.520 ± 34.422 pg/ml respectively (Figure 7B). At the 24th hour, the production of IL-6 by pre-treated cells with the same extract decreased slightly compared to that produced by untreated cells. Indeed, the average quantity of IL-6 produced by untreated cells is 367.500 ± 34.422 pg/ml. After a pretreatment with 50 and 100 μg/ml of the extract, we observed a decrease in IL-6 production by PBMCs with average concentrations of 312.915 ± 34.422 and 286.973 ± 34.422 pg/ml respectively (Figure 7C). PBMCs were or were not treated with 50 or 100 μg/ml of ethanolic extract from A. hispidum and for 30 min were stimulated with 10 μg/ml of LPS. The cultivations were collected after 4 h (A), 16 h (B) and 24 h (C). Rats IL-6 was dosed in these supernatants with the IL-6’s BioLegend commercial dosing kit (p < 0.01).
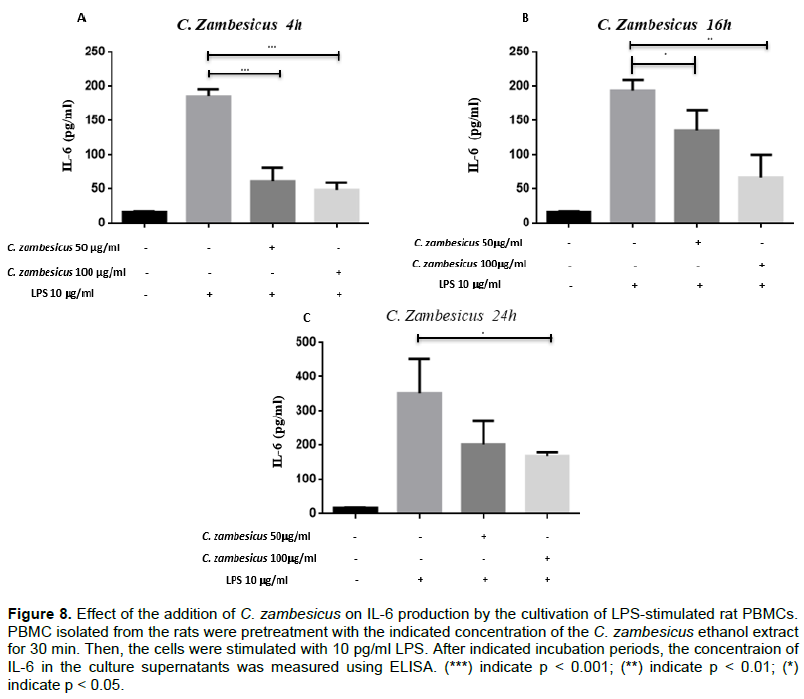
The ethanolic extract of C. zambesicus at 50 μg/ml or 100 μg/ml resulted in a decrease in the quantity of IL-6 secreted at 4 h compared to the quantity of IL-6 produced by cells that were not pretreated. The average quantity of IL-6 produced by untreated cells is 185.453 ± 34.422 pg/ml. After pretreatment with 50 and 100 μg/ml, we observed a decrease in IL-6 production by PBMCs with average concentrations of 61.993 ± 34.422 pg/ml (p = 0.001) and 48.560 ± 34.422 pg/ml (p = 0.000) respectively (Figure 8A). This decrease is also observed at the sixteenth hour. The average quantity of IL-6 produced by untreated cells at the 16th hour is 194.197 ± 34.422 pg/ml. After pretreatment with 50 and 100 μg/ml of C. zambesicus ethanolic extract, we observed a decrease in IL-6 production by PBMCs with average concentrations of 143.630 ± 42.158 pg/ml (p = 0.016) and 67.293 ± 34.422 pg/ml (p = 0.002) respectively (Figure 8B). At the 24th hour, the ethanolic extract of C. zambesicus at 50 and 100 μg/ml resulted in a decrease in the quantity of IL-6 secreted by PBMCs compared to the quantity of IL-6 produced by cells that were not pretreated. Indeed, the average quantity of IL-6 produced by untreated cells is 367.500 ± 42.158 pg/ml. After pretreatment with 50 and 100 μg/ml of C. zambesicus ethanolic extract, we observed a decrease in IL-6 production by PBMCs with average concentrations of 235.687 ± 34.422 and 173.265 ± 42.158 pg/ml (p = 0.023) respectively (Figure 8C). PBMCs were or were not treated with 50 or 100 μg/ml of C. zambesicus ethanolic extract for 30 min and stimulated with 10 μg/ml of LPS. The cultivations were collected after 4 h (A), 16 h (B) and 24 h (C). Rats IL-6 was dosed in these supernatants with the BioLegend commercial dosing kit of IL-6 (p < 0.01 and p < 0.001).
DISCUSSION
Phytochemical screening
Phytochemical screening of the leaves of A. hispidum revealed the presence of cathechic tannins, flavonoids, reductive compounds, mucilages, steroids and terpens, saponosides, coumarins and quinones. These results are consistent with those of Abubakar et al. (2015). However, these authors found the presence of alkaloids, which is contrary to this study. This could be due to a low concentration of alkaloids in the leaves of A. hispidum or to climatic conditions, to the soil quality of the two regions where the work was carried out. The work of N'do et al. (2018) also revealed the presence of phenolic compounds and flavonoids in the aqueous and ethanolic extracts of A. hispidum leaves. However, the results are identical to those of Pansambal et al. (2017) who found the tannins, flavonoids, steroids, terpenes, saponosides and coumarins in the extracts of the same plant. Roy et al. (2010) found carbohydrates, alkaloids, glycosides, flavonoids, tannins and saponosides. These results are in part consistent with the results of this study. The phytochemical study of C. zambesicus leaves revealed the presence of cachetic tannins, flavonoids, reductive compounds (sodium sulfite, sodium dithionite, sodium thiosulfate, and aldehydes), mucilages and coumarins. These results are on one hand consistent with the work of Reuben et al. (2009) who found in the acetate ethyl extract of C. zambesicus, terpens, steroids, reductive compounds and flavonoids. However, these authors did not look for mucilages and coumarins. In contrast the work of Mohamed et al. (2016) found in addition to the compounds found in this study, alkaloids, saponins and anthraquinones in the ethyl acetate extract of C. zambesicus. In addition, Ajayi and Omomagiowawi (2018) found glucosides, steroids, anthraquinones, phenols, saponosides and flavonoids in C. zambesicus ethanolic extract. These differences may be related to the nature of the solvent used or the assay method of the metabolites. Other authors have shown that these plants have useful purposes in traditional medicine (Yapi, 2015).
Larval cytotoxicity
The study of larval cytotoxicity of ethanolic extracts from both plants yielded LC50 greater than 0.1mg/ml (0.75mg/mL and 0.36mg/mL respectively for A. hispidum and C. zambesicus. Therefore, it is concluded that the ethanolic extracts of these two plants are not toxic to the larvae of Artemia salina. This result is consistent with that of Hougbeme et al. (2014) who found a LC50 of 0.41mg/mL for A. hispidum. As for C. zambesicus, we have not found earlier studies of larval toxicity by its extracts.
Anti-inflammatory activity
The results of anti-inflammatory activity showed that ethanolic extracts from the C. zambesicus and A. hispidium leaves were divergent to an increase in the volume of rat leg edema induced by formalin at 2%. These extracts therefore have an anti-inflammatory effect. This would justify their traditional use as medicinal plants in the treatment of inflammation. The anti-inflammatory effect of ethanolic extract (200 mg/kg) of both plants is observed already from the first hour and is maximum at the fourth hour of the five hours after injection. The anti-inflammatory effect of ethanolic extract from C. zambesicus roots was demonstrated in a study by Okokon and Nwafor (2010) through the reduction in volume of carrageenan-induced edema in the rat's paw. Compared to A. hispidum no studies in the literature have treated the anti-inflammatory activity of its extracts. However, several studies have shown antibacterial, antifungal and antiparasitic activity in extracts from both plants (Abubakar et al., 2015; Edewor and Olajire, 2011; Manal et al., 2015; Pansambal et al., 2017; Reuben et al., 2009). In addition, phytochemical screening of ethanolic extracts from both plants revealed the presence of phenolic compounds such as tannins and flavonoids that are thought to have anti-inflammatory activity (Chika et al., 2012; Duru et al., 2013; Enechi and Nwodo, 2014).
The anti-inflammatory activity of extracts from C. zambesicus and A. hispidum leaves may be due to flavonoids and tannins. Flavonoids, natural antioxidants, have been reported to play a very important role in the treatment of inflammation, tumours and bacterial diseases (Duru et al., 2013; Ezeja et al., 2011). Tannins are also endowed with antioxidant properties which leads to the activation of the immune defense and reduce the risk of cancer and degenerative diseases (Houmenou et al., 2018; Mpondo et al, 2012).
Immunomodulatory activity
In relation to immunomodulatory activity, the objective is to determine the action of ethanolic extracts of A. hispidum and C. zambesicus on the production of two pro-inflammatory cytokines (TNF-α and IL-6) by PBMCs of LPS-stimulated rats. The results showed that both extracts inhibit the production of IL-6 and only the ethanolic extract from the C. zambesicus leaves inhibited the production of TNF-α. Several studies showed the involvement of cytokines in inflammatory processes (Lotrich, 2012; Wojdasiewicz et al., 2014; Zhang and An, 2009). Anti-inflammatory drugs modulate the synthesis of inflammation mediators such as pro-inflammatory cytokines (Muster, 2005; Noack and Kolopp-Sarda, 2018). With most of these anti-inflammatory drugs with side effects (Guilpain and Le Jeunne, 2012), research is focused on the discovery of new plant-based products with fewer side effects (Garcia-Lafuente et al., 2009; Vikrant and Arya, 2011; Yatoo et al., 2018). These products, to be effective, must be able to act on mediators of inflammation including cytokines (Amro et al., 2013). This study of the anti-inflammatory activity of ethanol extracts of A. hispidum and C. zambesicus showed the ability to reduce the volume of edema induced by formalin at 2%. If these extracts have induced a decrease in edema, they could therefore act on the mediators of inflammation. This work evaluated the action of these extracts on the production of TNF-α and IL-6 by mononucleated peripheral blood cells (PBMCs) of rats, following stimulation by LPS.
The effect of the extracts on the production of IL-6 by PBMCs was sought in vivo by feeding rats by these extracts. To do this, the rats were fed 200 mg/kg of the ethanolic extract of A. hispidum and 5 ml of blood was collected from rats 4 h after feeding. Stimulation of their PBMCs by LPS showed an inhibition of IL-6 production. Oral feeding would have resulted in the extract being disseminated through the digestive tract and then through the bloodstream. This would reflect a systemic effect of the extract. Other authors have also shown that plant extracts act in vivo on inflammation mediators (Mukherjee et al., 2014; Noosud et al., 2017).
The results of our study showed that the ethanol extract of A. hispidum, at the doses used, had no significant effect on the production of TNF-α by PBMCs of rats stimulated by LPS. This could be related to the mechanisms of action of this extract or to the doses used (Diallo, 2019). Since this extract showed its action on edema (according to the results of our study on anti-inflammatory activity) without acting on the production of TNF-α, it could likely have local and non-peripheral actions.
On the other hand, the ethanol extract of C. zambesicus (100 μg/ml) led to a decrease in TNF-α production in LPS-stimulated PBMCs cultured supernatants (10 μg/ml). The extract may inhibit the activation of mononucleated cells of peripheral blood, producing TNF-α (Bradley, 2008). Indeed, during an inflammatory response, an increase in the biosynthesis of Toll Like Receptor (TLR) is observed (Cen et al., 2018). Studies have shown that LPS induces an increase in the level of expression of LRT-4 (Huet et al., 2004; Shukla et al., 2018). C. zambesicus ethanolic extract may inhibit the expression of these receptors by PBMCs or attach to these receptors to prevent the induction of TNF-α production (Bùfalo et al., 2014; Davicino et al., 2007; Zhao et al., 2020). It could also act on the signaling pathways leading to the production of TNF-α. The NF-kB (Nuclear-Factor-kappa B) track is one of the routes whose activation results in the production of TNF-α (Schottelius and Baldwin, 1999). The extract used could act on this path. Moreover, the TNF-α having a pleiotropic action (Noack and Miossec, 2017), its inhibition would have acted on other mediators of inflammation. The TNF-α's action also depends on its connection to its two receivers (TNFR1 and TNFR2) (Aggarwal, 2003; Bradley, 2008). TNF-α's binding to its TNFR1 receptor is conducive to the development of inflammation and tissue damage, while its binding to the TNFR2 receptor maintains local homeostasis and activates tissue regeneration (Kalliolias and Ivashkiv, 2016). In this context, the activity of the C. zambesicus ethanolic extract would promote the binding of TNF-α to TNFR2 receptors. In addition, TNF-α is synthesized as a precursor associated with the plasma membrane (Pro-TNF-α) and is released in soluble form by proteolytic cleavage under the action of a TACE enzyme (TNF-α-Converting Enzyme) (Canault et al., 2006). The extract could also block the action of this enzyme (Letavic et al., 2002; Schaal et al., 2018; Trifilieff et al., 2002).
We observed an important inhibition of IL-6 produced by the PBMCs of Wistars rats stimulated by LPS after pre-treatment of cells with ethanolic extracts of A. hispidum and C. zambesicus. The percentages of reduction range from 60.21 to 73.81%. Several mechanisms could be responsible for this process.
IL-6 is one of the key cytokines of inflammation (Noack and Miossec, 2017). The inhibition action of the extracts on this cytokine would confirm their anti-inflammatory properties. IL-6 is secreted by a large number of cells including T and B lymphocytes and monocytes (Cantagrel et al., 2017). The extracts could block the stimulation of these cells by LPS. In this case, the TLR-4 receptors expressed by these cells may have been blocked (Bùfalo et al., 2014; Davicino et al., 2007; Zhao et al., 2020).
Like other pleiotropic cytokines (Thu et al., 2014), inhibition of IL-6 production may influence the synthesis of other inflammation mediators. Indeed, IL-6 induces the synthesis of proteins of the acute phase of inflammation such as C-reactive protein (CRP), amyloid serum protein A, fibrinogen and hepcidine by hepatocytes (Tanaka et al., 2014). Extracts that inhibited the synthesis of IL-6 may also inhibit the synthesis of CRP, and other proteins in the acute phase of inflammation (Hollebeeck et al., 2012; Kometani et al., 2008; Makni et al., 2011; Parhiz et al., 2015).
Moreover, cell signaling leading to the inflammatory response is a complex phenomenon that involves several signaling pathways that interact with each other; cytokines operate in networks (Cavaillon, 1995) with autocrine, paracrine, synergistic, pleiotropic, cascading and antagonistic actions (Zhang and An, 2009). This means that the effect of the extracts on the signaling pathway of a cytokine, could lead to actions on other mediators. In this context, a molecule with an anti-inflammatory effect by acting on one or more mediators would mitigate the overall inflammatory response. This study showed that the extracts inhibited the production of two pro-inflammatory cytokines. If these extracts have anti-inflammatory effects, they could stimulate the production of other anti-inflammatory cytokines such as IL10, TGF-β, IL4 and IL13 (Cavaillon, 1995).
The different anti-inflammatory actions of the extracts of these plants could be related to the secondary metabolites they contain. In fact, phytochemical screening of the leaves of these two plants has shown the presence of flavonoids that may have an inhibitory effect on the production of pro-inflammatory cytokines (Kim et al., 2004; Kumar and Pandey, 2013; Middleton et al., 2000; Parhiz et al., 2015). Several other studies have also highlighted the anti-inflammatory effects of other polyphenols through their inhibitory actions on the production of inflammation mediators. In addition, these polyphenols also act on signaling pathways such as the NF-kB track and the MAPK track (Jang et al., 2012; Park et al., 2011).
The reducing edema and inhibition capabilities of the production of pro-inflammatory cytokines of these extracts show the anti-inflammatory potentials of these plants (Okokon et al., 2013; Okokon and Nwafor, 2010). These observations could justify the use of these plants by market herbalists in the treatment of inflammatory diseases. It is therefore imperative to further research in the direction of isolating the active compounds in order to think about the development of new anti-inflammatory drugs.
CONCLUSION
Traditionally, many medicinal plants are used for the treatment of different ailments. Pharmaco-toxicological tests are necessary to justify therapeutic uses and also to suggest a more effective means to enhance the use of medicinal plants. The presence of polyphenolic compounds in the extracts of A. hispidium and C. zambesicus, justifies the biological properties (antioxidant, anti-inflammatory, anti-infectious...) attributed to these non-cytotoxic plants by the herbalists of Benin. In addition, in vitro anti-inflammatory tests have shown that the ethanolic extracts of these two plants have the capacity to reduce oedemas. Thus, these two plants can be used to relieve patients suffering from an infection by reducing edema in the inflamed tissues due to the infection. Moreover, the ethanolic extracts of A. hispidium and C. zambesicus are able to inhibit the production of inflammation mediators such as the cytokines TNF-α and IL-6. Therefore, these two plants are then potential candidates for bio-guided studies to discover new molecules or to develop new anti-inflammatory drugs.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Abubakar A, Sodipo OA, Khan IZ, Fugu MB, Mamza UT, Gulani IA (2015). Phytochemical Screening and in-Vitro Antimicrobial Activities of the Leaf Extract of Acanthospermum hispidum DC (Asteraceae). Journal of Plant Studies 4(2):66-73. |
|
|
Agbankpé AJ, Bankolé SH, Assogba F, Dougnon TV, Yèhouénou B, Gbénou J, Baba-Moussa L (2015). Phytochemical Screening and Cytotoxic Analysis of Three Local Vegetables Used in the Treatment of Bacterial Diarrhoea in Southern Benin (West Africa): A Comparative Study. British Biotechnology Journal 9(4):1-13. |
|
|
Aggarwal BB (2003). Signalling pathways of the TNF superfamily: A double-edged sword. Nature Reviews. Immunology 3(9):745-756. |
|
|
Ambe ASA, Ouattara D, Tiebre M-S, Vroh BTA, Zirihi GN, N 'guessan KE (2015). Diversity of medicinal plants used in the traditional treatment of diarrhea in the markets of Abidjan (Ivory Coast). Journal of Animal &Plant Sciences 26(2):4081-4096. [Article in French]. |
|
|
Amoroso M, Rossol M, Heine H, Meusch U, Quandt D, Klein C, Sweet MJ, Hauschildt S (2012). LPS-induced cytokine production in human monocytes and macrophages. Critical Reviews in Immunology 31(5):379-446. |
|
|
Amro BI, Haddadin RN, Tawaha K, Mohammad M, Mashallah S, Assaf AM (2013). In vitro antimicrobial and anti-inflammatory activity of Jordanian plant extracts: A potential target therapy for Acne vulgaris. African Journal of Pharmacy and Pharmacology 7(29):2087-2099. |
|
|
Ajayi AO, Omomagiowawi IE (2018). Antimicrobial Activity of Croton zambesicus on Staphylococcus aureus and Streptococcus species. SYLWAN 160(10):114-124. |
|
|
Block S, Baccelli C, Tinant B, Van Meervelt L, Rozenberg R, Jiwan JLH, Llabrès G, De Pauw-Gillet MC, Quetin-Leclercq J (2004). Diterpenes from the leaves of Croton zambesicus. Phytochemistry 65(8):1165-1171. |
|
|
Bradley JR (2008). TNF-mediated inflammatory disease. The Journal of Pathology 214(2):149-160. |
|
|
Bruneton J (1999). Pharmacognosie - Phytochimie, plantes médicinales. Lavoisier, Paris. p. 1120. |
|
|
Búfalo MC, Bordon-Graciani AP, Conti BJ, De Assis Golim M, Sforcin JM (2014). The immunomodulatory effect of propolis on receptors expression, cytokine production and fungicidal activity of human monocytes. The Journal of Pharmacy and Pharmacology 66(10):1497-1504. |
|
|
Canault M, Peiretti F, Kopp F, Bonardo B, Bonzi MF, Coudeyre JC, Alessi MC, Juhan-Vague I, Nalbone G (2006). The TNF alpha converting enzyme (TACE/ADAM17) is expressed in the atherosclerotic lesions of apolipoprotein E-deficient mice: Possible contribution to elevated plasma levels of soluble TNF alpha receptors. Atherosclerosis 187(1):82-91. |
|
|
Cantagrel A, Degboé Y, Constantin A, Davignon JL (2017). TNF-?, interleukin-6 and interleukin-1: 3 pivotal cytokines in rheumatoid arthritis pathogenesis. Revue Du Rhumatisme Monographies 84(4):325-330. |
|
|
Cavaillon JM (1995). Inflammation: a precarious balance between pro- and anti-inflammatory cytokines. Revue Française des Laboratoire 1995(276):27-35. |
|
|
Cen X, Liu S, Cheng K (2018). The role of toll-like receptor in inflammation and tumor immunity. Frontiers in Pharmacology 9(878):1-8. |
|
|
Chika E, Moses I, Chigozie U, Olisa A, Ifeanyichukwu I, Michael A, Charles E (2012). Evaluation of antibacterial activity of the leave extracts of Buchholzia coriacea. Asian Journal of Pharmaceutical and Biological Research 2(4):204-208. |
|
|
Davicino R, Mattar A, Casali Y, Porporatto C, Correa SG, Micalizzi B (2007). In vivo immunomodulatory effects of aqueous extracts of Larrea divaricata Cav. Immunopharmacology and Immunotoxicology 29(3-4):351-366. |
|
|
Deguenon E, Dougnon T, Sènou M, Agbankpé J, Hounmanou Y, Bankolé H, Dokpomiwa HA, Dougnon J, Baba-Moussa L (2017). Biological and chemical activities of some beninese plant's extracts. International Journal of Biology, Pharmacy and Allied Sciences 6(12):2333-2358. |
|
|
Diallo I (2019). Antioxidant and anti-inflammatory potentials of Lentinula edodes (Shiitake) sporophores under different culture conditions. Human medicine and pathology, University of Montpellier: French ffNNT:2019MONTG042ff. fftel-02481192. [document in French]. |
|
|
Dougnon TV, Bankolé HS, Edorh AP, Dougnon TJ, Klotoé JR, Loko F, Boko M (2013). Cytotoxicity of Leaves and Fruits of Solanum macrocarpon Linn ( Solanaceae ) against shrimp larvae ( Artemia salina Leach ). Research Journal of Recent Sciences 2(5):6-9. |
|
|
Duru M, Ugbogu A, Amadi B, Odika P, Anudike OCJ, Osuocha K (2013). Chemical constituents of Buchholzia coriacea seed. Proceedings of the 35th Annual International Conference: Workshop & Exhibition of Chemical Society of Nigeria 2:39-45. |
|
|
Edewor TI, Olajire AA (2011). Two flavones from Acanthospermum hispidum DC and Their Antibacterial Activity. International Journal of Organic Chemistry 1(3):132-141. |
|
|
Enechi OC, Nwodo OFC (2014). Anti-ulcer and gastric anti-secretory activities of seed extract of Buchholzia coriacea in Wistar Albino rats. African Journal of Biotechnology 13(27):2755-2761. |
|
|
Ezeja MI, Ezeigbo II, Madubuike KG (2011). Analgesic activity of the methanolic seed extract of Buchholzia coriacea. Research Journal of Pharmaceutical, Biological and Chemical Sciences 2(1):187-193. |
|
|
García-Lafuente A, Guillamón E, Villares A, Rostagno MA, and Martínez JA (2009). Flavonoids as anti-inflammatory agents: implications in cancer and cardiovascular disease. Inflammation Research 8(9):537-552. |
|
|
Guedje NM, Tadjouteu F, Dongmo RF, Fokunang CN, Fotso S (2012). Traditional African medicine (TAM) and phytomedicines: challenges and development strategies. Health Sciences and Diseases 12(3):1-25. |
|
|
Guilpain P, Le Jeunne C (2012). Anti-inflammatory and immunosuppressive effects of glucocorticoids. La Presse Medicale 41(4):378-383. |
|
|
Hollebeeck S, Winand J, Hérent MF, During A, Leclercq J, Larondelle Y, Schneider YJ (2012). Anti-inflammatory effects of pomegranate (Punica granatum L.) husk ellagitannins in Caco-2 cells, an in vitro model of human intestine. Food and Function 3(8):875-885. |
|
|
Houmenou V, Adjatin A, Assogba F, Gbenou J, Akoegninou A (2018). Phytochemical and Cytotoxicity Study of Some Plants Used In The Treatment Of Sterility In Southern Benin. European Scientific Journal February 14(6):156-171. |
|
|
Houngbeme AG, Gandonou C, Yehouenou B, Kpoviessi SDS, Sohounhloue D, Moudachirou M, Gbaguidi FA (2014). Phytochemical Analysis, Toxicity and Antibacterial Activity of Benin Medicinal Plants Extracts Used in the Treatment of Sexually Transmitted Infections Associated With Hiv/Aids. International Journal of Pharmaceutical Sciences and Research 5(5):1739-1745. |
|
|
Huet O, Choukroun G, Mira JP (2004). Toll like receptor, inflammatory response and sepsis. Reanimation 13:167-175. |
|
|
Jang EY, Yang CH, Han MH, Choi YH, Hwang M (2012). Sauchinone suppresses lipopolysaccharide-induced inflammatory responses through Akt signaling in BV2 cells. International Immunopharmacology 14(2) :188-194. |
|
|
Kalliolias GD, Ivashkiv LB (2016). TNF biology, pathogenic mechanisms and emerging therapeutic strategies. Nature Reviews: Rheumatology 12(1):49-62. |
|
|
Kim HP, Son KH, Chang HW, Kang SS (2004). Anti-inflammatory plant flavonoids and cellular action mechanisms. Journal of Pharmacological Sciences 96(3):229-245. |
|
|
Klavina L (2018). Composition of mosses, their metabolites and environmental stress impacts. Doctoral in Biology and Environmental Science, University of Latvia. 130 pp. |
|
|
Klavina L, Springe G, Nikolajeva V, Martsinkevich I, Nakurte I, Dzabijeva D, Steinberga I (2015). Chemical composition analysis, antimicrobial activity and cytotoxicity screening of moss extracts (moss phytochemistry). Molecules 20(9):17221-43. |
|
|
Kometani T, Fukuda T, Kakuma T, Kawaguchi K, Tamura W, Kumazawa,Y, Nagata K (2008). Effects of α-glucosylhesperidin, a bioactive food material, on collagen-induced arthritis in mice and rheumatoid arthritis in humans. Immunopharmacology and Immunotoxicology 30(1) :117-134. |
|
|
Kpodji P, Lozès E, Dougnon V, Assogba P, Koudokpon H, Baba-Moussa L (2019). Utilisation des plantes du sud-bénin dans le traitement des maladies inflammatoires?: enquête ethnopharmacologique auprès des herboristes. Revue Ivoirienne des Sciences et Technologie 34:127-143. |
|
|
Kumar S, Pandey AK (2013). Chemistry and Biological Activities of Flavonoids?: An overview. The Scientific World Journal, ID 162750:1-17. |
|
|
Letavic MA, Axt MZ, Barberia JT, Carty TJ, Danley DE, Geoghegan KF, Halim NS, Hoth LR, Kamath AV, Laird ER, Lopresti-Morrow LL, McClure KF, Mitchell PG, Natarajan V, Noe MC, Pandit J, Reeves L, Schulte GK, Snow SL, Sweeney FJ, Tan DH, Yu CH (2002). Synthesis and biological activity of selective pipecolic acid-based TNF-α converting enzyme (TACE) inhibitors. Bioorganic and Medical Chemistry Letters 12(10):1387-1390. |
|
|
Lotrich F (2012). Inflammatory Cytokines, Growth Factors, and Depression. Current Pharmaceutical Design 18(36):5920-5935. |
|
|
Luni? TM, Oal?e MM, Mandi? MR, Sabovljevi? AD, Sabovljevi? MS, Gaši? UM, Duleti?-Lauševi? SN, Boži? BD, Boži? Nedeljkovi? BD (2020). Extracts Characterization and In Vitro Evaluation of Potential Immunomodulatory Activities of the Moss Hypnum cupressiforme Hedw. Molecules 25(15):3343. |
|
|
Makni M, Chtourou Y, Fetoui H, Garoui EM, Boudawara T, Zeghal N (2011). Evaluation of the antioxidant, anti-inflammatory and hepatoprotective properties of vanillin in carbon tetrachloride-treated rats. European Journal of Pharmacology 668(1-2):133-139. |
|
|
Manal AI, Amna MA, Mona AM, Nafesa AN, Sulafa HH (2015). Antimicrobial and antioxidant activity of seed extracts of Croton zambesicus. Scholars Journal of Applied Medical Sciences 3(7C):2665-2667. |
|
|
Mandi? MR, Oal?e MM, Luni? TM, Sabovljevi? AD, Sabovljevi? MS, Gaši? UM, Duleti?-Lauševi? SN, Boži? BD, Boži? Nedeljkovi? BD (2021). Chemical characterization and in vitro immunomodulatory effects of different extracts of moss Hedwigia ciliata (Hedw.) P. Beauv. from the Vrša?ke Planine Mts., Serbia. PLoS ONE 16(2): e0246810. |
|
|
Middleton E, Kandaswami C, Theoharides TC (2000). The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharmacology Reviews 52(4):673-751. |
|
|
Mohamed NA, Amna A, Ahmed SK (2016). In vitro antioxidant activity and phytochemical screening of Croton zambesicus. Journal of Pharmacognosy and Phytochemistry 12(56) :12-16. |
|
|
Mousseux M (1995). Toxicity test on Artemia salina larvae and maintenance of a debalane farm. French Pacific University, University Centre of New Caledonia: France. p. 21. [document in french]. |
|
|
Mpondo ME, Dibong SD, Ladoh YCF, Priso RJ, Ngoye A (2012). Phenol plants used by the populations of the city of Douala. Journal of Animal and Plant Sciences 15:2083-2098. [Article in french] |
|
|
Mukherjee PK, Nema NK, Bhadra S, Mukherjee D, Braga FC, Matsabisa MG (2014). Immunomodulatory leads from medicinal plants. Indian Journal of Traditional Knowledge 13(2):235-256. |
|
|
Muster D (2005). Anti-inflammatory drugs. EMC-Stomatologie 1 :21-29. |
|
|
Noack M, Kolopp-Sarda M-N (2018). Cytokines and inflammation: physiology, pathophysiology and therapeutic use. Revue Francophone des Laboratoire 499:28-37. |
|
|
Noack M, Miossec P (2017). Selected cytokine pathways in rheumatoid arthritis. Seminars in Immunopathology 39(4):365-383. |
|
|
Noosud J, Lailerd N, Kayan A, Boonkaewwan C (2017). In vitro and in vivo assessment of inhibitory effect of stevioside on pro-inflammatory cytokines. Avicenna Journal in phytomedicine 7(2):101-107. |
|
|
Okokon JE, Nwafor PA (2010). Antiinflammatory, analgesic and antipyretic activities of ethanolic root extract of Croton zambesicus. Pakistan Journal of Pharmaceutical Sciences 23(4):385-392. |
|
|
Okokon JE, Nwafor PA, Ekpo MD (2010). Subchronic toxicity studies of the ethanolic root extract of Croton zambesicus. Pakistan Journal of Pharmaceutical Sciences 23(2):160-169. |
|
|
Ouahchia C, Cherif H-S, Hamaidi-Chergui F, Marzen L, Deradji S, Hemma R, Nouar N, Saidi F (2017). Acute and subacute toxicity of methanolic extracts of Inula viscosa (Dittrichia viscosa L.). Revue Agrobiologia, 7(2):562-573. |
|
|
Pansambal S, Deshmukh K, Savale A, Ghotekar S, Pardeshi O, Jain G, Aher Y, Pore D (2017). Phytosynthesis and biological activities of fluorescent CuO nanoparticles using Acanthospermum hispidum L. extract. Journal of Nanostructures, 7(3):165-174. |
|
|
Parhiz H, Roohbakhsh A, Soltani F, Rezaee R, Iranshahi M (2015). Antioxidant and anti-inflammatory properties of the citrus flavonoids hesperidin and hesperetin: An updated review of their molecular mechanisms and experimental models. Phytoherapy Research 29(3):323-331. |
|
|
Park BS, Lee JO (2013). Recognition of lipopolysaccharide pattern by TLR4 complexes. Experimental and Molecular Medecine 45(12):e66-9. |
|
|
Park SE, Sapkota K, Kim S, Kim H, Kim SJ (2011). Kaempferol acts through mitogen-activated protein kinases and protein kinase B/AKT to elicit protection in a model of neuroinflammation in BV2 microglial cells. British Journal of Pharmacology 164(3):1008-1025. |
|
|
Petrovska BB (2012). Historical review of medicinal plants' usage. Pharmacognosy Reviews 6(11):1-5. |
|
|
Reuben KD, Fanna IA, Akan JC, Olufunke AS (2009). Phytochemical screening and antimicrobial studies of ethyl acetate extract of Croton zambesicus Muell Arg. Stem Bark. The Pacific Journal of Science and Technology 10(2):842-849. |
|
|
Roy H, Chakraborty A, Bhanja S, Shankar N, Sruti R, Mishra B, Ellaiah P (2010). Preliminary phytochemical investigation and anthelmintic activity of Acanthospermum hispidum DC. Journal of Pharmaceutical Science and Technology 2(5):217?221. |
|
|
Salatino A, Salatino M, Negri G (2007). Traditional uses, Chemistry and Pharmacology of Croton species (Euphorbiaceae). Journal of the Brazilian Chemical Society 18(1):11-33. |
|
|
Schaal JB, Maretzky T, Tran DQ, Tran PA, Tongaonkar P, Blobel CP, Ouellette AJ, Selsted ME (2018). Macrocyclic θ-defensins suppress tumor necrosis factor-α (TNF-α) shedding by inhibition of TNF-α-converting enzyme. Journal of Biological Chemistry 293(8):2725-2734. |
|
|
Schottelius A, Baldwin J (1999). A role for transcription factor NF- kB in intestinal inflammation. International Journal of Colorectal Disease 14(1):18-28. |
|
|
Shukla R, Ghoshal U, Ranjan P, Ghoshal UC (2018). Expression of toll-like receptors, pro-, and antiinflammatory cytokines in relation to gut microbiota in irritable bowel syndrome: The evidence for its micro-organic basis. Journal of Neurogastroenterology and Motility 24(4):628-642. |
|
|
Tanaka T, Narazaki M, Kishimoto T (2014). IL-6 in Inflammation, Immunity, and Disease. Cold Spring Harbor Perspectives in Biology, 6(10):1?16.Thu HLT, Van ANT, Thanh HT, Dieu TNT, Minh HLT (2014). Role of cytokines in asthma. Journal Of Functional Ventilation and Pulmonology 5(14):25-32. |
|
|
Trifilieff A, Walker C, Keller T, Kottirsch G, Neumann U (2002). Pharmacological profile of PKF242-484 and PKF241-466, novel dual inhibitors of TNF-α converting enzyme and matrix metalloproteinases, in models of airway inflammation. British Journal of Pharmacology 135(7):1655-1664. |
|
|
Vikrant A, Arya ML (2011). A review on anti-inflammatory plant barks. International Journal of PharmTech Research 3(2):899-908. |
|
|
World Health Organization (WHO) (2002). WHO Strategy for Traditional Medicine 2002-2005 World Health Organization, Geneva p.1-78. [document in french]. |
|
|
Winter CA, Risley EA, Nuss GW (1962). Carrageenin-induced edema in hind paw of the rat as an assay for antiiflammatory drugs. Proceedings of the Society for Experimental Biolody and Medicine. Society of Experimental Biology and Medicine 111(3):544-547. |
|
|
Wojdasiewicz P, Poniatowski LA, Szukiewicz D (2014). The role of inflammatory and anti-inflammatory cytokines in the pathogenesis of osteoarthritis. Mediators Inflammation, 2014. 561459 |
|
|
Xie CF, Lou HX (2009). Secondary metabolites in bryophytes: an ecological aspect. Chemistry & biodiversity 6(3):303-312. |
|
|
Yapi BA, Gnahoue G, Camara D, Zirihi G (2015). Botanical study, evaluation of the Antifungal Activity of the leaves of Acanthospermum Hispidum Dc, (Asteraceae) on the in vitro growth of Candida albicans and toxicity study on human HFF cells. European Scientific Journal 11(30):225-237. [Article in french] |
|
|
Yatoo MI, Gopalakrishnan A, Saxena A, Parray OR, Tufani NA, Chakraborty S, Tiwari R, Dhama K, Iqbal HMN (2018). Anti-inflammatory drugs and herbs with special emphasis on herbal medicines for countering inflammatory diseases and disorders - A Review. Recent Patents on Inflammation and Allergy Drug Discovery 12(1):39-58. |
|
|
Zhang J-M, An J (2007). Cytokines, Inflammation and Pain. International Anesthesiology. Clinics 45(2):27-37. |
|
|
Zhao L, Li M, Sun K, Su S, Geng T, Sun H (2020). Hippophae rhamnoides polysaccharides protect IPEC-J2 cells from LPS-induced inflammation, apoptosis and barrier dysfunction in vitro via inhibiting TLR4/NF-κB signaling pathway. International Journal of Biological and Macromolecules 155:1202-1215. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


