Full Length Research Paper

ABSTRACT
Tea Tree oil (TTO), the essential oil from the Australian native Melaleuca alternifolia has demonstrated a variety of beneficial efficacies including antimicrobial, antifungal, antiviral and anti-inflammatory. This report discusses data obtained on the in vitro activity of TTO on human glioblastoma cells U87MG. Cell viability was examined by 2-(4,5-dimethyl-2-thiazolyl)-3,5-diphenyl-2H-tetrazolium bromide MTT assay. Growth was investigated by incubating cells with various concentrations of TTO (0.025 and 0.05 %) for 24, 48 or 72 h and daily cell count. Cell cycle and apoptosis assay were assessed by flow citometry. TTO decreased cell viability in a dose and time-dependent manner. . The cell cycle distribution showed that TTO enhanced the accumulation of the cells in G0/G1 phase. The analysis by Western blot of protein related to cell cycle (CDK2 and p27), cell apoptosis (caspase 6 and 8), necrosis (TNFR1 and RIP1) demonstrated that TTO induces U87MG growth inhibition by more synergic mechanisms: necrosis, low level apoptosis and cell cycle arrest. TTO induces also in vivo glioblastoma tumor growth inhibition in a murine subcutaneous model.
Key words: Brain cancer, tea tree oil, terpinol, glioblastoma, natural drug, adjuvant chemotherapy, temozolomide, apoptosis, cell cycle.
INTRODUCTION
Glioblastoma (GBM) is the most common primary malignant form of brain cancer, with a median survival of 7-15 months from the time of diagnosis. Hallmarks of the aggressive cancer include extensive infiltration and strong vascular proliferation into the surrounding brain parenchyma (Wei et al., 2014; Kim et al., 2015; Van-Tellingen et al., 2015). GBM is characterized by a pronounced mitotic activity, high neoangiogenesis, cellular heterogeneity, necrosis and high proliferative rates (Cicero et al., 2013; De-Almeida et al., 2012). Furthermore, the presence of cancer stem cells, was able to proliferate and generate glial neoplastic cell, contribute to the unfavourable prognosis of GBM patients (Surawicz et al., 1998). Conventional therapy for glioblastoma, tumor resection followed by radiotherapy and chemotherapy typically temozolomide (TMZ), is limited in efficacy due to high rates of recurrence, overall resistance to therapy, and devastating neurological deterioration (Kim et al., 2015; Lin et al., 2015). Even though numerous efforts have been made to identify the molecular pathways and potential “druggable” targets involved in gliomagenesis (Shergalis et al., 2018), gross-total resection of tumor tissue, followed by adjuvant chemo- and radiotherapy remain the standard of care (Stupp and Weber, 2005). Current therapeutic approaches to glioblastoma are often associated with the development of multidrug resistance, important side effects, and high cost, underscoring the unmet need for more efficacious and less toxic interventions. Numerous experimental data and other clinical-pathological studies have shown that overexpression of the xenobiotic transporter P-glycoprotein and MGMT not methylated lead to drug-resistant phenotype, often associated with more aggressive behavior glioma (Diao et al., 2019; Sarkaria et al., 2008). Resistance to cancer agents through drug metabolism, drug target mutation (Gottesman, 2002), compartmentalization (Kitazono et al., 1999) and through primary resistance; such as over-2 expression of MDR1 (the gene for P-glycoprotein) and especially apoptotic gene mutations of cancer cells, which render them resistant to apoptosis, means the search for novel antitumour agents that may overcome this is warranted. Tea tree oil (TTO), the essential oil from the Australian native Melaleuca alternifolia has demonstrated a variety of beneficial efficacies including antimicrobial (Carson et al., 2006; Akthar et al., 2014), antifungal (Hammer et al., 2003), antiviral (Schnitzler et al., 2001) and anti-inflammatory (Hart et al., 2000). TTO consists of over 100 components; the most represented are: terpinen-4-ol, γ-terpinene, α-terpinene and 1,8- cineole. It is the most abundant component, terpinen-4-ol that is the likely mediator of the in vitro and in vivo efficacy TTO (Mondello et al., 2006). Amongst the activities listed above, anti-cancerous efficacy has also been identified (Gautam et al., 2014). A single study demonstrated terpinen-4-ol and TTO in vitro anticancer activity. Human melanoma and Adriamycin resistant melanoma cells treated with TTO and terpinen-4-ol underwent caspase dependent apoptosis, a process thought to involve plasma membrane interaction via lipid reorganisation (Calcabrini et al., 2004;Tawona et al., 2014). Interestingly, both TTO and terpinen-4-ol were more effective against the resistant cell line suggesting that neither are substrates for P-glycoprotein (P-gp), a very useful property in the treatment of multidrug resistant (MDR) tumours. Only one study has demonstrated in vitro anticancer efficacy of 1,8-cineole against two human leukemia cells lines through apoptosis (Moteki et al., 2002). Accordingly, further study of the potential anticancer activity of TTO and its major components is warranted. Currently, no investigation of TTO or terpinen-4-ol has been conducted in vivo and before in vitro data can be translated clinically, in vivo studies must be completed. In this study we investigate the effect of TTO and components in vitro coupled with an investigation of the effects of TTO on tumour development in immuno-competent murine tumour models. Studies on human melanoma cells have shown that both oil and its main active component, terpinen-4-ol, are able to induce caspase-dependent apoptosis (Calcabrini et al., 2004). Other research has shown that tea tree oil has in vitro antitumor activity on breast cancer cells, causing a cell cycle arrest in the S phase (Assmann et al., 2018). The vegetal kingdom has always represented an attractive source for therapeutics and several examples do exist for natural products being included in current protocols to tackle the limits of chemotherapy. One of the main objectives of brain cancer research is therefore to identify new effective therapeutic strategies against drug-resistant tumors, in fact the increase in resistance and the side effects of current therapeutic approaches are leading scientists to explore alternative medicines to the traditional one as an option for finding new chemical entities for cancer treatment (Gautam et al., 2014). Among phytochemicals, essential oils have been considered attractive for their wide variety of bioactivities. Anticancer potential of essential oils has been explored and several studies are now available in the literature. Traditionally, essential oils have been used for their biological activities including antiseptic, analgesic, sedative, anti-inflammatory, spasmolytic, and locally anesthetic properties (Sharifi et al., 2017). Furthermore, they are used in aromatherapy for health improvement due to their sedative or stimulant properties (Hadfield et al., 2001). Essential oils (also called volatile or ethereal oils) are aromatic, highly volatile, hydrophobic liquids produced by aromatic plants as secondary metabolites. Essential oils (EO) have been the subject of research for alternative medicine research; EOs-mediated therapy cannot replace standard chemotherapy and radiotherapy but can be used in combination with cancer therapy to reduce drug side effects (Gautam et al., 2014). On the basis of their chemical structures the constituents of essential oils are classified as terpene hydrocarbons, divided into monoterpenes (C10), sesquiterpenes (C15) and diterpenes (C20); terpenes containing oxygen (terpenoids), such as alcohols, ketones, aldehydes, esters, lactones, coumarins and phenylpropanoids. Among the phytochemicals, essential oils were considered interesting for their wide variety of bioactivity (Bozin et al., 2006). The anti-tumor potential of oils has been explored and numerous studies are now available in the literature. Traditionally, EOs are used for their biological activities including antiseptic, analgesic, sedative, anti-inflammatory, spasmolytic and local anesthetic properties (Javad et al., ,2017). Furthermore, they are used in aromatherapy for health improvement and especially to reduce the agony of patients with brain cancer (Hadfield et al., 2001). It has been suggested that topical application of aromatic oils may exert antibacterial, anti-inflammatory and analgesic effects. For cancer patients, the benefits obtained include reduced levels of anxiety and relief from emotional stress, pain, muscle tension and fatigue (Fellowes et al., 2004). There are a lot of evidence that essential oils and their constituents act according to multiple pathways and mechanisms that involve apoptosis, cell cycle arrest, antimetastatic and antiangiogenic effect (Gautam et al., 2014). Tea Tree Oil is an essential oil, derived from the M. alternifolia plant, used largely for its antimicrobial properties and incorporated as an active ingredient in many topical formulations used for the treatment of skin diseases (Carson et al., 2006). It is difficult to identify the single component responsible for each activity of Tea Tree Oil as the oil, as shown in Table 1, consists of a mix of substances. The antitumor activity is carried out above all by the terpene fraction which is the one present in greatest concentration. Studies on human melanoma cells have shown that both oil and its main active component, terpinen-4-ol, are able to induce caspase-dependent apoptosis (Calcabrini et al., 2004). Other research has shown that tea tree oil has in vitro antitumor activity on breast cancer cells, causing a cell cycle arrest in the S phase (Assmann et al., 2018). The ability to inhibit the growth of cancer cells has been shown in a dose-dependent manner in cancer of the colon, pancreas, prostate and stomach (Shapira et al., 2016). The main purpose of the present study was to investigate the effects of TTO on glioma cells both in vitro and in vivo and to explore the possible anticancer action and mechanisms that lead TTO-induced apoptosis , cell cycle arrest and necrosis.
MATERIALS AND METHODS
Cell culture
For in vitro and in vivo experiments we used a human glioblastoma continuous cell line U87MG, from Sigma Aldrich Collection (LGC Promochem, Teddington, UK). Glioblastoma cells were grown in Dulbecco’s Modified Eagle’s Medium (Gibco) supplemented with 10 % fetal bovine serum, 2 mmol/L-glutamine, 50 U/mLl Penicillin and 50 µg/mLl Streptomycin, at Streptomycin, at 37°C, 5 % CO2, and 95 % of humidity. For treatment, we used Tea Tree Oil 38 % (ESI and Temozolomide (Sigma Aldrich).
Growth inhibition of U87MG by TTO treatment
To evaluate, in vitro, the response to treatment with TTO, human glioblastoma cells (U87MG) were plated in 48-well plates, 1×104 cells per well, in Dulbecco 's Modified Eagle' s Medium (DMEM, Invitrogen) supplemented with 10 % fetal calf serum (Invitrogen) and incubated at 37 °C in an atmosphere containing 5 % CO2. The day after the cells were treated with TTO 0.025 and 0.05 % TTO. The substance used is pure TTO 38 %, produced by ESI srl Registered office Corso Ferrari 74/6 17011 Albisola Superiore (SV) Italy Tax Code: 01099380105 VAT number: 11264680155 REA number: SV-102611.
For the experiment, the TTO was reconstituted in 0.5% dimethylsulfoxide (DMSO) and Tween 80 (0.002%) (Bag and Chattopadhyay, 2015). At 24, 48 and 72 hrs after treatment we made a cell count by Burker chamber, in the presence of trypan blue dye (Sigma) to discriminate live cells from dead ones. The same experiment was repeated in presence of canonical chemotherapic TMZ. Cells were seeded in 48-well plates (1×104 cells/well) and were treated for 24, 48, and 72 hrs with 0.025 and 0.05 % TTO alone or in combination with 10 µM TMZ. At the end of the treatment, cell count was performed.
Cell viability assays
U87MG cells were seeded in 96-well plates (5x103 cells/well) in DMEM 10 % FBS and allowed to adhere for 12 hrs. Cells were exposed to different concentrations of TTO (0.025 and 0.05 %) for 24, 48 and 72 hou. After treatment, 5 mg/mLl MTT (Methylthiazolyldiphenyl-tetrazolium bromide; Sigma–Aldrich) in 100 µl of DMEM without phenol red was added to the cells for 2 hrs. We washed cells with PBS (phosphate-buffered saline) and added 100 µl of DMSO. MTT metabolized concentrations was determined by spectrophotometer measurement of absorbance at 570 nm.
Analysis of cell cycle, apoptosis and necroptosis proteins after treatment with TTO
We analyzed cell cycle, apoptosis and necroptosis proteins in U87Mg control and U87Mg after short and long treatment with TTO 0.025 %. In short-term treatments we plated U87Mg (800,000 per plate) in DMEM without serum for 48 hrs. After re-addition of FBS we carried out inductions with TTO 0.025 % for 2, 4, 8, 12 and 24 h. For long-term treatments, we plated U87Mg in DMEM 10 % FBS and we induced with TTO 0.025 % only on the first day or daily for 24 rs, 48 rs and 72 hrs. Proteins were extracted from U87MG in Triton X-100 lysis buffer (Tris-HCL 10 mM, EDTA 1 mM, NaCl 150 mM, Triton X-100 1 %, NaF 1 mM, 1 mM Na4P2O7, 1 mM Na3VO4, protease inhibitors 1X). Proteins (40 μg) were separated by SDS-PAGE on a polyacrylamide gel and transferred to nitrocellulose membranes (Amersham) by electroblotting. The membranes were incubated for 2 h at room temperature in 5 % milk or BSA diluted in T-TBS 1X, and then incubated over night at 4°C with primary antibodies specific and 1 h with secondary antibody. Each membrane was incubated with mouse monoclonal anti-βactin (1:50000; Santa Cruz Biotechnology). Protein were detected by chemiluminescence using ECL Western blotting (Amersham). The signals were detected by a digital scanner and quantified by densitometric analysis (Scion Image software). For cell cycle proteins analysis we used anti- CDK2, anti-P21 and anti-P27 primary antibodies (1:1000), all from Cell Cycle Regulation Sampler Kit (Cell Signaling Technology); we also analysed P53 protein with mouse monoclonal antibody anti-P53 (Roche Diagnostic). For the analysis of the proteins involved in apoptosis we used anti-Caspase 6, anti-Caspase 8 and anti-Parp primary antibodies (1:1000) from the Procaspase Antibody Sampler Kit (Cell Signaling technology), while for necroptotic proteins we used anti-TNFR1, anti-RIP and anti-TRADD primary antibodies (1:1000), all from Death Receptor antibody sampler Kit (Cell Signaling technology); as secondary antibodies we used an anti-rabbit or anti-mouse (1: 2000; Cell Signaling technology).
FACS analysis by flow cytometry
The U87MG human GBM cells were plated (8 × 105) in DMEM with 10 % FBS and treated with TTO (0.025 %) for 24, 48 and 72 hrs. After treatment, cells were trypsinized, washed in sample buffer (glucose 0.1 % in HBSS), fixed in 70 % ethanol, and stored at 4 °C over night until the day of analysis. Before analysis, Propidium iodide (50 μg/mLl) was added for 30 min at room temperature. Flow Cytometry (FC) analysis of the cell cycle was performed with Gallios instrument (Beckman Coulter).
Apoptosis assay
U87MG were treated with TTO 0.025 % for 24, 48 and 72 h. Cells were collected for apoptosis assay with Annexin V-FITC apoptosis detection kit (Cell Signaling). After centrifugation at 1200 rpm for 5 min, the pellet (5 x 105 cells) was resuspended in 1ml of 1 x binding buffer with 5 µl of Annexin V-FITC conjugated and 10 µl of Propidium iodide solution. The reaction complex was incubated for 10 minutes on ice in the dark. Samples were subjected to apoptosis analysis assay by flow cytometry. The expression of Annexin V was tested also by immunofluorescence on U87MG treated with TTO 0.025 % for 24 hrs.
U87MG cell implantation
Male nude mice CD1 (Charles River, Calco, CO, Italy; 20-22 g body weight) were housed in a controlled conditions (temperature 22 °C; humidity, 40 %) on a 12 hrs light/dark cycle and observed daily. The experiments were carried out according to the regulations for the care and use of animals promulgated by National Institutes of Health. After anesthesia with ketamine 100 mg/Kg and xyilazine 10 mg/Kg i.p., mice were subcutaneously implanted with 5 × 106 U87MG cells/0.2 mLl (matrigel and DMEM). At 7 days after inoculation treatments are carried out with TTO 3.5 % intratumoral, twice a week, for 3 weeks.The dose administered in vivo was calculated according to evidence in the literature (Shapira et al., 2016).Tumor size was measured by caliper. Subsequently, the mice were sacrificed by cervical dislocation after anesthesia, and tumors removed, were fixed in 4 % formalin, dehydrated in ethanol at increasing concentrations and embedded in paraffin. From each tumors were cut sections of 4 μm thickness. The sections were stained with Mayer's hemalum (Diapath) and Eosyn (Diapath) and subjected to microscopic evaluation.
Immunofluorescence on tumor tissue
Tissue sections of 4 µm thickness were deparaffinized in xylene, rehydrated in alcohol with decreasing concentration and permeabilized in Triton 0.1% for 30 min. We performed an incubation with 10 % goat serum for 1 h and then with primary antibody anti-TNFR1 (1: 100, Cell Signaling) over night at 4°C. Secondary antibody anti rabbit -Cy3 was added (1: 200 Vector) for 1 h and DAPI mounting medium (Vector). The slides have been analysed by a fluorescence microscope at 40x magnification.
Apoptosis detection of tumor tissue by TUNEL staining
Tissue sections of 4 µm thickness were deparaffinized in xylene, rehydrated in alcohol with decreasing concentration and fixed in 4% methanol. TUNEL tests were performed by a commercial kit, Deadend fluorometric Tunel System (Promega) following the manufacturer's instructions. The slides were counterstained with DAPI and analysed by a fluorescence microscope at 40x magnification.
Statistical analysis
Statistical analysis was performed using statistical software (GraphPad Prism Software, version 5.0). Values are presented as the Mean±S.E.M. Student's t-test was used to compare two groups. Analysis of variance (one-way ANOVA) was used to compare three and more conditions and Dunnett post-test was used to compare mean values for all groups. Differences between groups were considered statistically significant when the null hypothesis pH0 was than P< 0.05.
RESULTS
Effect of TTO on the of human GBM cells growth
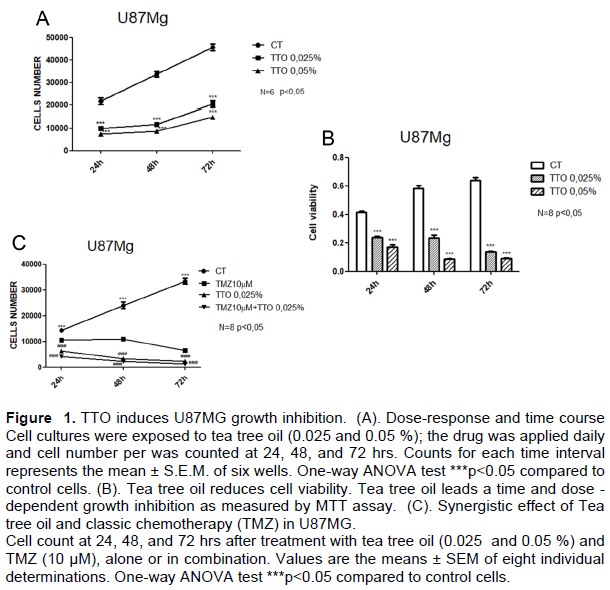
The effect of TTO on GBM cell (U87MG) proliferation was analyzed long term, with cell count performed at 24, 48, and 72 hrs after TTO treatment. As shown in Figure 1 Hyperlink reference not valid.A, the continuous cell line U87MG had a significant reduction in proliferation with 0.025 and 0.05 % TTO; a more significant reduction appears at 72 hrs of treatment. The TTO concentration that caused the greatest growth inhibition in cell line U87MG was 0.05 %. The growth inhibition is dose and time dependent.
TTO inhibits the viability of U87MG glioblastoma cells
To determine the effect of TTO on the cell viability of U87MG glioblastoma cells, U87MG were treated at various concentrations of TTO 0.025, 0.05% for different time (24, 48 and 72 hrs). Cells with 0.1 % DMSO were used as a control. The MTT assay showed that TTO inhibited the viability of U87MG cells in a time and dose-dependent manner (Figure 1B). TTO concentrations 0.05% markedly suppressed the cell viability of U87MG cells (PP < 0.005).
TTO treatment amplified the effects of TMZ on the proliferation of human glioma cells in vitro
The continuous cell line U87MG were treated with TMZ (10 μM) alone or in combination with TTO 0.025%, and after 24, 48, and 72 h, the activity of the drugs on cell count was evaluated. The TMZ treatment appeared to be effective, but a statistically significant reduction in growth was observed in cells treated with a combination of the 2 drugs (10 μM TMZ plus 0.025% TTO) already after 24 h of treatment (Figure 1C).
Western blot analysis
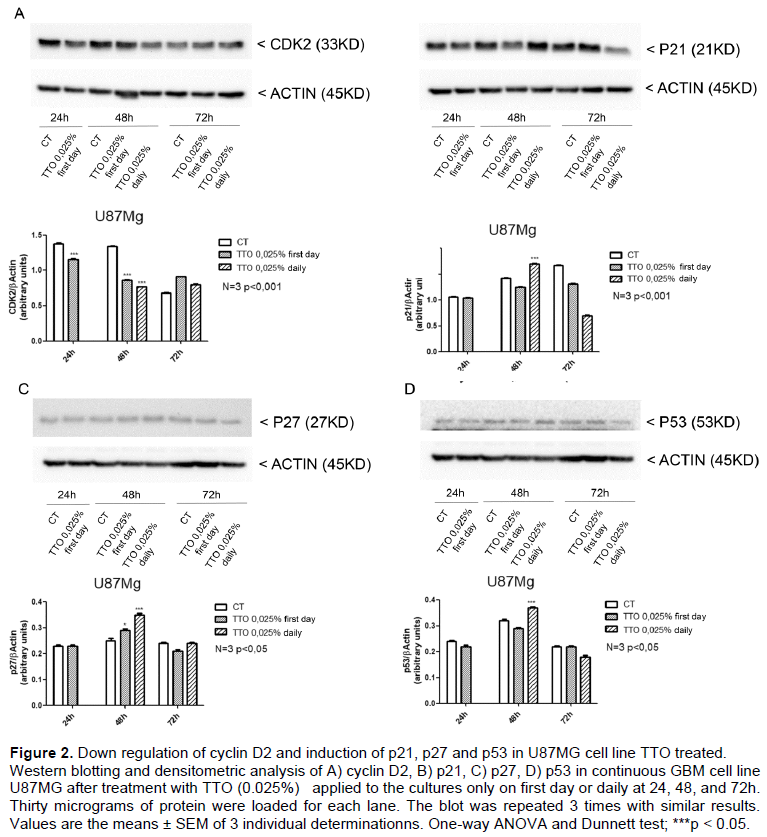
To assess the molecular mechanism of TT0 -induced cell-cycle arrest, we investigated the expression levels of cell cycle related proteins in U87MG control and TTO -treated. TTO treatment (0.025%) causes a reduction of cyclin D2 in U87MG cells at 2, 4, 8 and 12 h (Supplementary Figure 1) of treatment and at long term exposure (24 and 48 h) (Figure 2A) while p21 levels was induced significatively only at 48 h by TTO treatment (0.025%) (Figure 2B). Cyclin-dependent kinase inhibitor p27 was also investigated, western blot analysis shows an increase of p27 at 48 h from TTO induction (Figure 2C). Finally, we analyzed the expression of the p53 oncogene that undergoes a significant increase at 48 h of treatment with TTO, p53 would participate in TTO-induced glioblastoma growth inhibition (Figure 2D).
TTO leads apoptotic effect in U87MG
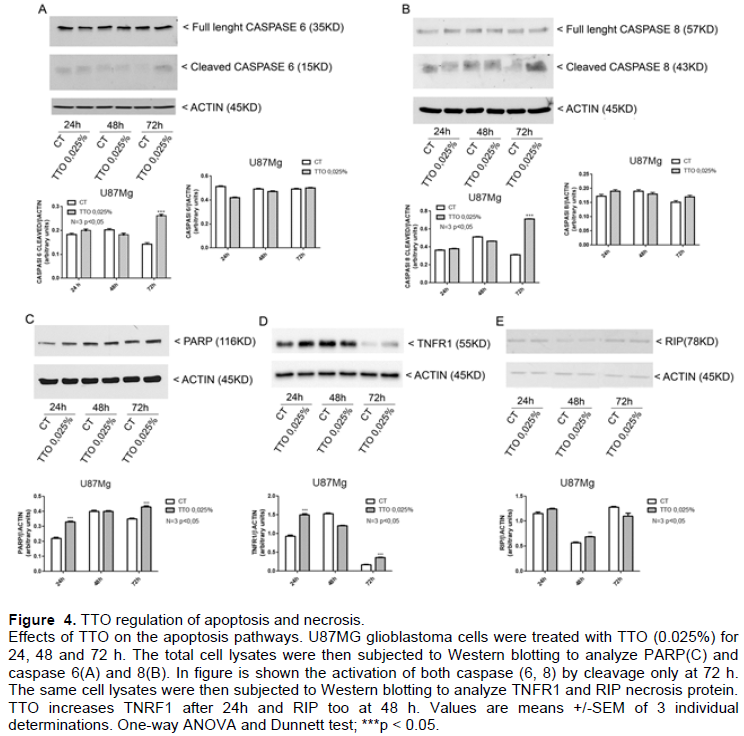
To investigate the pro-apoptotic effect of TTO in U87MG glioblastoma cells PARP expression was analyzed by western blot on U87MG TTO treated with 0.025% for 24, 48 and 72 h. The expression of PARP (Figure 4C) reveals in the cells treated with TTO an accumulation of PARP (116Kd) and cannot be seen of cleavage form.
Since the cleavage of PARP protein occurs only in case of apoptosis, increased expression of normal form in cells treated with TTO suggest the switch to other mechanism than control growth inhibition and Necrosis. The expression of caspase 6 and caspase 8 was also analyzed; Figure 4A and B shows that there is an increase of cleavage caspase 6 and caspase 8 after 72 h TTO treatment, witnessing the presence of a real apoptotic process also.
TTO induces necrosis in U87MG
We study the protein that normally induces inflammation TNFR1 by western blot of cell lysate of U87MG treated with 0.025% TTO for 24, 48 and 72 h. The results showed an increase of TNFR1 at 24 h while RIP at long term exposure of TTO (48 h) (Figures 4D and E) and increase of TNFR1 was found just at short treatment times (2, 4 and 8 h) whereas TRADD increase only at 4 rshrs of treatment (Supplementary Figure 1). The increase in these proteins suggests the coexistence of a phenomenon of necrosis.
TTO induces G0/G1 cell cycle arrest in U87MG cells
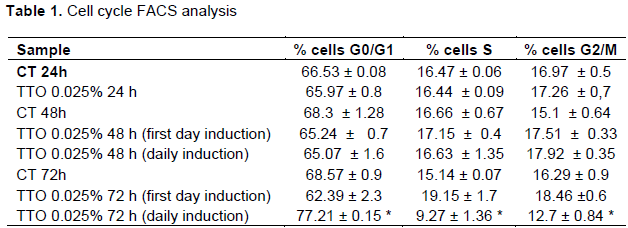
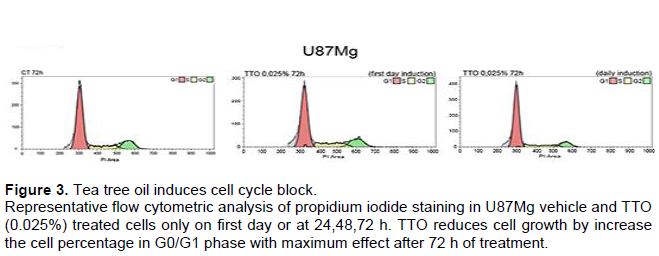
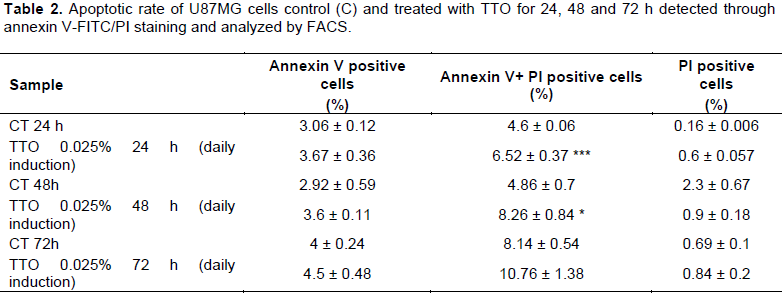
To explore how TTO inhibits the proliferation of U87MG cells, FACS was performed to determine cell cycle distribution. After 24 h plating U87MG were treated with 0.025% of TTO for 24, 48, and 72 h. After 72 h of treatment is visible the maxim effect on cell cycle arrest, the percentage of U87MG cells staying at G0/G1 phase were 68.57± 0.9 in the control while in treated cell is 77± 0.15 and 37.9 ± 1.1 (Figure 3). The increase of G0/G1 phase cell population was accompanied by a concomitant decrease in % cells in S and G2 phase (Figure 3). Our results indicated that TTO triggers cell cycle arrest in the G0 / G1 phase in a time and dose dependent manner showing a significant effect after 72 h. To demonstrate the presence of apoptosis in TTO-treated cells a FACS experiment with Annexin V was performed. The results of the analysis show that with increasing treatment time: (24, 48 and 72 h) the peak relative to cells is increased in late apoptosis phase. (Table 2). Annexin V immunofluorescence reveals an increase of phosphatidylserine externalisation in TTO treated cells (supplementary Figure 2).
In vivo experiment
We implanted U87MG cells subcutaneously onto right side of nude mice (N8), after one week from implantation, mice were divided randomly in two groups: control and TTO treated one. Mice were treated with TTO 3.5% by intratumoral injectionintratumoral injection twice for week for 3 weeks. At the end of the treatments animals were sacrificed and tumors taken from subcutaneous were weighed and undergoing to histological analysis. Tumor size was evaluated weekly by caliper. The evaluation of tumor size, by caliper measurements analysis confirm that, in mice treated with TTO, tumor size significantlysize significantly reduced about 80% (Figure 5C). In Figure 5D there is a representative graph of tumor growth that is visibly inhibited massively by treatment with TTO.
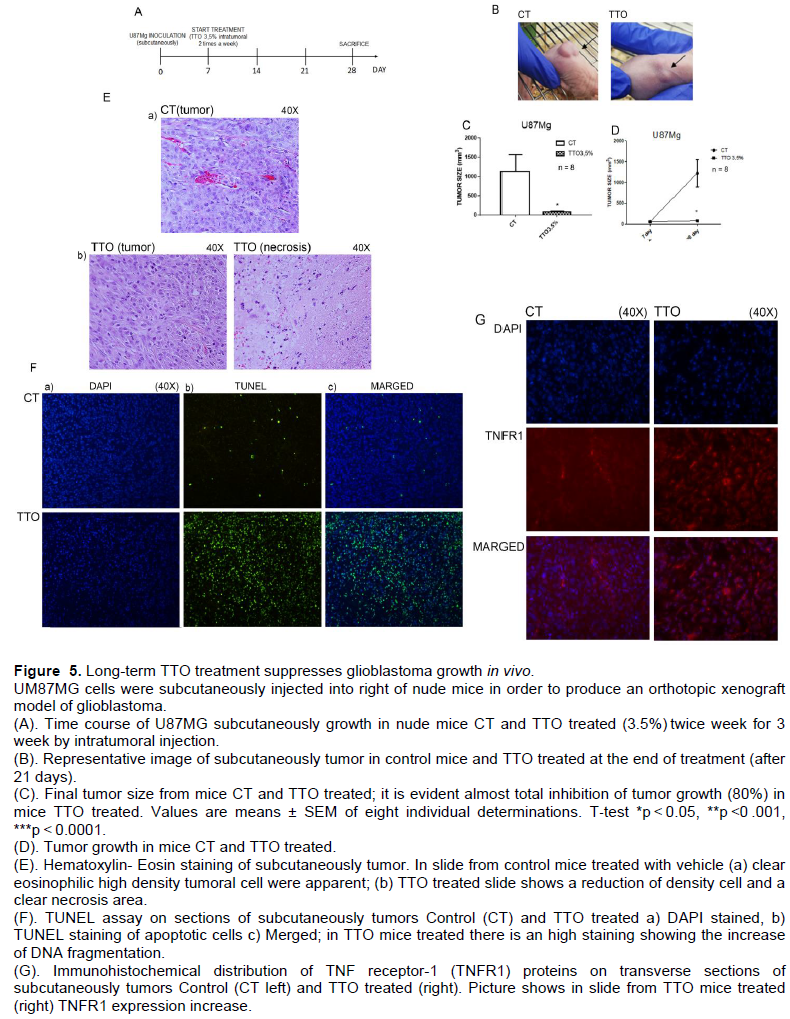
Cytologically, tumors were composed of large pleomorphic cells with abundant eosinophilic cytoplasm (Figure 5E). Hematoxylin -Eosin staining shows that tumors are composed by dysmorphic cells, typical of glioblastoma, with high cell density; the tumors treated with TTO show the same cell morphology but a marked reduction in cell density with large areas of necrosis. Finally, to show that even in the tissues there is an evidence of pro-apoptotic effect of TTO we exposed the same slide to TUNEL assay.
TUNEL assay and immunofluorescence of pro-inflammatory receptor TNRF1
To consider the possibility that TTO induced apoptosis in vivo, paraffin-embedded sections of tumors were assessment of apoptosis by TUNEL staining. In the control group, the structure of cells was intact and the nucleus was completely stained blue. However, in the TTO -treated group, the percentage of TUNEL-positive cells (green) markedly increased (Figure 5F), confirming the increase in apoptotic tumor cells of DNA fragmentation after TTO treatment. On the same sections we analyzed by Immunofluorescence the pro-inflammatory receptor TNFR1 as shown in (Figure 5G) TNFR1 expression increase visible in tumor slide from TTO treated.
DISCUSSION
Despite standard treatments for glioblastoma (GMB), relapse and drug resistance remain the main causes of treatment failure in many patients.
Glioblastoma multiforme (GBM), which is a Grade IV brain tumor according to the World Health Organization (WHO) classification, is the most common form of primary brain tumor in the central nervous system (CNS), and its aggressive nature and evasiveness to treatments make it one of the most lethal cancers. Less than 3% of glioblastoma patients are still alive at 5 years after diagnosis. (Ohgaki et al., 2005).
Current treatments for GBM range from common chemotherapeutic agents such as temozolomide (TMZ) in conjunction with radiotherapy (Stupp protocol 23) (Stupp and Weber, 2005). Over the past 10 years, however, therapeutic agents have not significantly increased the median survival of patients with glioblastoma. The 5- year survival rate for patients with the same disease, after treatment including surgical resection, radiotherapy and chemotherapy is less than 9.8 %. In order to improve the results of brain tumor therapy, in our laboratory we investigated the use of natural and less toxic substances that can be used as adjuvant drugs in the treatment of glioblastoma. Complementary medicines with natural compounds (Vengoji et al., 2018; Arcella et al., 2015 Kamarudin et al, 2019; Correa et al, 2019) have become increasingly popular in recent decades. In this study we investigated about the effect Tea Tree oil, an essential oil that comes from an Australian plant, M. alternifolia, made up of different substances including terpinen4-ol, considered the active component, which, as shown by other studies has anticancer effect, is able to induce a significant inhibition of pancreatic, prostatic, gastric and colorectal cancer tumor cells (Nakayama et al., 2017; He et al, 2000). In this study, for the first time, the effect of TTO on human GMB cell line was studied, both in vitro and in vivo. An in vitro proliferation test was performed on U87MGg cells to evaluate their proliferation in the presence of different concentrations of TTO (0.025 and 0.05 %). Both doses tested caused a growth inhibition of about 50 %, a block confirmed also by the MTT toxicity test. The inhibition of TTO on glioblastoma cell growth was tested also in combination of TTO with canonical chemoterapic for brain tumors temozolomide, the results of cell count and MTT assay shown an interestingly synergic effect. At this point we tried to investigate the expression of proteins involved in apoptosis, necrosis and control of the cell cycle to establish with which mechanism the growth block occurs. Western blot analysis showed that, in cells treated with TTO for 2, 4, 8, 12 and 24 hrs CDK2 protein decrease significatively while the tumoroncosuppressor p27 increase only at 8 h from TTO treatment. We found also an increase of caspase. 8 and caspase 6 cleavage, and p53 increases. These results suggest that the cause of the growth inhibition of U87MG glioblastoma cells by TTO is a results of more synergic mechanism: may be an increase of apoptosis and a block of the cell cycle in G0/G1 phase. This hypothesis was confirmed by FACS analysis: longterm treatment 72 h with TTO in U87MG caused an increase of cells in the G0/G1 phase.
Apoptosis and cell blocking therefore would not represent the unique and preponderant cause of TTO cell growth inhibition, as it is not massive apoptosis so it is conceivable that other mechanisms can act synergistic cooperating in blocking the cell growth. It could occur a phenomenon of necroptosis, for which we analyzed the expression of TNFR1 and TRADD which represents the most widely studied model of programmed necrosis (Justyna et al., 2014; Bing et al. 2018). The results showed an increase of TNFR1 and RIP at long term exposition of TTO (24, 48 and 72 hrs) and an increase of TNFR1 and TRADD at short treatment times (2, 4, 8 and and 12 hrs) which reveals that treatment with Tea Tree Oil could also lead to necroptosis. Necroptosis is a pro-inflammatory cell death in which swelling of the organelles, rupture of the cell membrane and uncontrolled release of intracellular contents into the surrounding tissue occurs. Necroptosis is a regulated necrosis mediated by death receptors (Berghe et al., 2014). It is well known that involvement of receptor like Fas, TNF, and TRAIL can lead to cell death through the recruitment of caspase-8 leading to initiation of extrinsic apoptotic pathway (Taylor et al., 2008). Considering the evidence of our results (increase of cleavage caspase 8, TNFR1, RIP, after 3 days TTO treatment) we have assumed necroptosis as the probably molecular mechanisms of necroptosis to death U87 Mg by TTO treatments. In contrast to apoptosis, necroptosis is caspase independent. These data together suggest that the growth block of U87MG, after TTO treatment, is determined by several control mechanisms: apoptosis, necrosis and cell blockade in G0/G1 phase. The cell line, U87MG, which has an amazing growth rate, replicates every 24 h, logarithmic growth, would explain the typical virulence of the brain tumor. The fact that a natural substance like TTO can block the growth of glioblastoma has comforted us to start an in vivo study in a more physiological context. In the in vivo studies, mice were subcutaneously injected with human glioblastoma U87MG, after 1 week were divided in two groups: control and treated one with TTO 3.5% (intratumoral administration two days a week for three weeks). Mice treated with TTO showed a 80% reduction in the tumor mass compared with tumor in control mice, as evidenced by volume. The histological analysis of the slide of removed tumors reveals typical hyperchromic pleomorphic elements; in the section from mice treated with TTO it can be evident in a big area of necrosis confirming that necrosis is one of the mechanisms underlying the growth block of glioblastoma by TTO. The presence of necrosis mechanism was confirmed by TUNEL assay too; in the section by tumor TTO treated for 3 weeks, as shown in the images, it is visible increase of DNA fragmentation.
CONCLUSION
A growing body of scientific reports has recently focused on the potential of essential oils as anticancer treatment in the attempt to overcome the development of multidrug resistance and important side effects associated with the antitumor drugs currently used (Russo et al, 2015). Our results indicated that TTO, the essential oil isolated from Melaleuca Alternifolia, is a natural compound that possesses the activity of inhibiting glioblastoma cell growth in vitro and in vivo at a dose- and time-dependent manner, and the mechanisms were associated with cell cycle arrest, triggering DNA damage and inducing apoptosis and necrosis. It is tempting to speculate that TTO might be utilized as a potential therapeutic agent against brain tumor. This is only a preliminary study to prove the antitumor effect of TTO on human glioblastoma cells.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
Dedicated to my father. We thank Rosanna de Rosa (English teacher) for the assistance rendered to edit the manuscript. The authors are also grateful to Dr. Vittorio Maglione (IRCCS Neuromed) for processing the Annexin V samples at the FACS.
REFERENCES
|
Akthar MS, Degaga B, Azam T (2014). Antimicrobial activity of essential oils extracted from medicinal plants against the pathogenic microorganisms. A review Issues in Biological Sciences and Pharmaceutical Research 2(1):001-007. |
|
|
Arcella A, Oliva MA, Staffieri S, Aalberti S, Grillea G, Madonna M, Bartolo M, Pavone L, Giangaspero F, Cantore G, Frati A (2015). In vitro and in vivo effect of human lactoferrin on glioblastoma growth. Journal of Neurosurgery 123(4):1026-1035. |
|
|
Assmann CE, Cadoná FC, Bonadiman BDSR, Dornelles EB, Trevisan G, Cruz IBMD (2018). Tea tree oil presents in vitro antitumor activity on breast cancer cells without cytotoxic effects on fibroblasts and on peripheral blood mononuclear. Biomedicine and Pharmacotherapy 103:1253-1261. |
|
|
Bag A, Chattopadhyay RR (2015). Evaluation of synergistic antibacterial and antioxidant efficacy of essential oils of spices in herbs. PLoS ONE 10(7):1-17. |
|
|
Berghe TV, Linkermann A, Jouan-Lanhouet S, Walczak H, Vandenabeele P (2014). Regulated necrosis: the expanding network of non-apoptotic cell death pathways. Nature Reviews Molecular Cell Biology 15:135-147. |
|
|
Bing S, Heling P, Ayaz N, Junying Y (2018). Necroptosis in development and diseases. Genes and Development 32(5-6):327-340. |
|
|
Bozin B, Mimica-Dukic N, Simin N, Anackov GJ (2006). Characterization of the volatile composition of essential oils of some lamiaceae spices and the antimicrobial and antioxidant activities of the entire oils. Journal of Agricultural and Food Chemistry 54(5):1822-1828. |
|
|
Carson CF, Hammer KA, Riley TV (2006). Melaleuca alternifolia (Tea Tree) Oil: a Review of Antimicrobial and Other Medicinal Properties. Clinical Microbiology Reviews 19(1):50-62. |
|
|
Calcabrini A, Stringaro A, Toccacieli L, Meschini S., Marra M., Colone M., Salvatore G, Mondellow F, Arancia G, Molinari A (2004). Terpinen-4-ol, The Main Component of Melaleuca Alternifolia (Tea Tree) Oil Inhibits the In Vitro Growth of Human Melanoma Cells. Journal of Investigative Dermatology 122:349 -360. |
|
|
Cicero C, Bonelli M, Mastrantoni E, Niccolini C, Laurenza M, Larocca LM, Pallini R, Traficante A, Spinsanti P, Ricci-Vitiani L, Arcella A, De Maria R, Nicoletti F, Battaglia G, Melchiorri D (2013). Type-3 metabotropic glutamate receptors regulate chemoresistance in glioma stem cells, and their levels are inversely related to survival in patients with malignant gliomas. Cell Death and Differentiation 20(3):396-407. |
|
|
Corrêa ACNTF, Vericimo MA, Dashevskiy A, Pereira PR, Paschoalin |
|
|
VMF (2019). Liposomal Taro Lectin Nanocapsules Control Human Glioblastoma and Mammary Adenocarcinoma. Cell Proliferation 24(3):471-477. |
|
|
De Almeida SF, Lunardi BA, Schwartsmann G, Roesler R, Abujamra AL (2012). Glioma revisited: from neurogenesis and cancer stem cells to the epigenetic regulation of the niche. Journal of Oncology 537861. |
|
|
Diao W, Tong X, Yang C, Zhang F, Bao C, Chen H, Liu L, Li M, Ye F, Fan Q, Wang J, Ou-Yang ZC (2019). Behaviors of Glioblastoma Cells in in Vitro Microenvironment. Scientific Report 9(1):85. |
|
|
Gautam N, Mantha AK, Mittal S (2014). Essential oils and their constituents as anticancer agents: a mechanistic view. BioMed Research International, Volume 2014, Article ID 154106, 23 pages. |
|
|
Gottesman MM (2002). Mechanisms of cancer drug resistance. Annual Review of Medicine 53: 615-627. |
|
|
Hammer KA, Carson CF, Riley TV (2003). Antifungal activity of the components of Melaleuca alternifolia (tea tree) oil. Journal of Applied Microbiology 95:853-860. |
|
|
Hadfield N (2001). The role of aromatherapy massage in reducing anxiety in patients with malignant brain tumours. International Journal of Palliative Nursing 7(6):279-285. |
|
|
Hart PH, Brand C, Carson CF, Riley TV, Prager RH, Finlay-Jones JJ (2000). Terpinen4-ol, the main component of the essential oil of Melaleuca alternifolia (tea tree oil), suppresses inflammatory mediator production by activated human monocytes. Inflammation Research 49(11):619-626. |
|
|
He J, Xiao Y, Zhang L (2000). In vitro cytotoxicity of Australian tea tree oil using human cell lines, Journal of essential oil research 9:575-582. |
|
|
Justyna S, Susann V, Sabine M, Arne L, Lutz T, Parvin D, Thomas H, Andreas L, Andrea R, Francis K M C, Dieter K, Stefan S, Dieter Adam (2014). TNF-induced necroptosis and PARP-1-mediated necrosis represent distinct routes to programmed necrotic cell death. Cellular and Molecular Life Sciences 71(2): 331-348. |
|
|
Kim SS, Harford JB, Pirollo KF, Chang EH (2015). Effective treatment of glioblastoma requires crossing the blood-brain barrier and targeting tumors including cancer stem cells: The promise of nanomedicine. Biochemical and Biophysical Research Communication 468:485-489. |
|
|
Kitazono M, Sumizawa T, Takebayashi Y, Chen ZS, Furukawa T, Nagayama S, Tani A, Takao S, Aikou T, Akiyama S (1999). Multidrug resistance and the lung resistance related protein in human colon carcinoma SW-620 cells. Journal of the National Cancer Institute 91:1647-1653. |
|
|
Lin KW, Liao A, Qutub AA (2015). Simulation predicts IGFBP2-HIF1α interaction drives glioblastoma growth. PLOS Computational Biology 11:e1004169 |
|
|
Mondello F, De Berbardis F, Girolamo A, Cassone A, Salvatore G (2006). In vivo activity of terpinen-4-ol, the main bioactive component of Melaleuca alternifolia Cheel (tea tree) oil against azole-suscetible and resistant human pathogenic Candida species. BMC Infectious Diseases 6:158. |
|
|
Moteki H, Hibasami H, Yamada Y, Katsuzaki H, Imai K, Komiya T (2002). Specific induction of apoptosis by 1,8-cineole in two human leukemia cell lines, but not an in human stomach cancer cell line. Oncology Reports 9:757-760. |
|
|
Nakayama K, Murata S, Ito H, Iwasaki K, Villareal MO, Zheng YW, Matsui H, Isoda H, Ohkohchi N (2017). Terpinen-4ol inhibits colorectal cancer growth via reactive oxygen species. Oncololy Letters 14(2):2015-2024. |
|
|
Ohgaki H, Kleihues P (2005). Epidemiology and etiology of gliomas. Acta Neuropathology 109 (1): 93-108. |
|
|
Russo R, Corasaniti MT, Bagetta G, Morrone LA (2015). Exploitation of Cytotoxicity of Some Essential Oils for Translation in Cancer Therapy. Evidence-Based Complementary and Alternative Medicine Volume 2015, Article ID 397821, 9 p. |
|
|
Sarkaria JN, Kitange GJ, James CD, Plummer R, Calvert H, Weller M, Wick W (2008). Mechanisms of Chemoresistance to alkylating agents in Malignant Glioma. Clinical Cancer Research 14(10):2900-2908. |
|
|
Schnitzler P, Schon K, Reichling J (2001). Antiviral activity of Australian tea tree oil and eucalyptus oil against herpes simplex virus in cell culture. Pharmazie 56:343-347. |
|
|
Shapira S, Pleban S, Kazanov D, Tirosh P, Arber N (2016). Terpinenol:A Novel and Promising Therapeutic Agent for Human Gastrointestinal Cancers. PLoS One 11(6):1-13. |
|
|
Sharifi-Rad J, Antoni Sureda A, Tenore GC, Daglia M, Sharifi-Rad M, Valussi M, Tundis R, Sharifi-Rad M, Loizzo MR, Ademiluyi A, Sharifi-Rad R, Ayatollahi SA, Iriti M (2017). Biological Activities of Essential Oils: From Plant Chemoecology to Traditional Healing Systems. Molecules 22(1):70. |
|
|
Shergalis A, Bankhead A, Luesakul U, Muangsin N, Nouri N (2018). Current Challenges and Opportunities in Treating Glioblastoma. Pharmacological Reviews 70(3):412-445. |
|
|
Stupp R, Weber DC (2005). The role of radio- and chemotherapy in glioblastoma. Onkologie 28(6-7):315-317. |
|
|
Surawicz TS, Davis F, Freels S, Laws ER Jr, Menck HR (1998). Brain tumor survival: results from the National Cancer Data Base. Journal Neurooncololy 40(2):151-6 |
|
|
Taylor RC, Cullen SP, Martin SJ (2008). Apoptosis: controlled demolition at the cellular level. Nature Reviews Molecular Cell Biology 9:231-241. |
|
|
Tawona NC, Du Plessis L, Gerber M, Hamman J, Du Plessis J (2014). Review of natural compounds for potential skin cancer treatment. Molecules 19(8):11679-11721. |
|
|
Van Tellingen O, Yetkin-Arik B, de Gooijer MC, Wesseling P, Wurdinger T, de Vries HE (2015). Overcoming the blood-brain tumor barrier for effective glioblastoma treatment. Drug Resistance Updates 19:1-12. |
|
|
Vengoji R, Macha MA, Batra SK, Shonka NA (2018). Natural products: a hope for glioblastoma patients. Oncotarget 9(31):22194-22219. |
|
|
Wei X, Chen X, Ying M, Lu W (2014). Brain tumor-targeted drug delivery strategies. Acta Pharmacologica Sinica B 4(3):193-201. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


