ABSTRACT
Experiments were carried out to investigate the egg-laying behaviour of the maize weevil, Sitophilus zeamais Motsch. (Coleoptera: Curculionidae) in uninfested and previously infested (termed conditioned here) maize grains. The number of offspring produced and preferred grain type of the weevil were examined under laboratory conditions, along with the effect of the presence of egg plugs on the oviposition site selection. The number of progeny produced was higher in conditioned maize. The highest and lowest number of adults emerged from conditioned and uninfested maize, respectively. S. zeamais individuals were significantly more attracted to conditioned grains compared to uninfested grains. S. zeamais females laid more eggs on host grains already bearing conspecific eggs (that is, conditioned) compared to clean grains (that is, uninfested). The number of eggs per grain showed a contagious distribution because females laid additional eggs on grains already infested with more than one egg. The results of this study indicate the possible role of the aggregation pheromone, intraspecific competition and egg marking in the observed egg-laying behaviour pattern of this pest species.
Key words: Maize weevil, egg, aggregation pheromone, oviposition behaviour, host marker.
In some stored agricultural products, adult female insect pests must select suitable grain on which to place their eggs because the larvae are not able to migrate to alternative grain sources. Consequently, insect larvae that develop and feed on the grain kernels of stored agricultural products are subjected to unique evolutionary problems. Adult females are able to vary their fitness according to whether they (1) space eggs out and minimise competition between larvae, or (2) lay eggs at random and risk losses in both the quantity and quality of offspring (Giga and Smith, 1985). The critical question is whether females are able to detect eggs that are already present within the seeds of host products.
For emerging larvae to receive an adequate food supply, several seed-oriented beetle species (Bruchidae) avoid ovipositing on host seeds that already bear conspecific eggs. For instance, Callosobruchus spp. use oviposition marker pheromones to regulate the distribution of eggs on seeds, to prevent the overcrowding of eggs on a few seeds, which would lead to intraspecific competition (Messina and Renwick, 1985a, b; Messina et al., 1987; Credland and Wright, 1990; MBata, 1992). In contrast, Sitophilus spp. eggs laid on the grains of stored agricultural products exhibit both random and aggregated distributions (Smith, 1986; Nardon et al., 1988).
Fava and Springhetti (1991) and Fava and Burlando (1995) observed that the females of the granary weevil, Sitophilus granarius L., lay more than one egg on the same grain, leaving other grains untouched. The authors concluded that the grain was not marked with pheromones during egg deposition because females did not appear to distinguish between infested and uninfested grains. Moreover, the egg distribution of the rice weevil, Sitophilus oryzae L., on grains fits the negative binomial distribution well, which is characterised by the fact that most egg plug distributions are underdispersed compared to the Poisson distribution (Holloway, 1984; Nardon et al., 1988). These examples show that different weevil species exhibit various oviposition strategies. Consequently, there is debate as to whether the genus Sitophilus oviposits at random or contagiously.
Thus, we investigated the egg-laying behaviour of S. zeamais, in an environment containing both uninfested maize and maize previously infested by S. zeamais. This study specifically aimed to determine (1) how adults distributed eggs between uninfested and infested (termed conditioned here) maize kernels, (2) the number of eggs laid and (3) the number of offspring produced in the two kernel types.
Grain and insects
Maize weevils were reared on shelled maize (Zea mays L.) for several generations under laboratory conditions. All insects (4-8 weeks old) were obtained from cultures maintained in the laboratory in darkened incubators at 30 ± 2°C and 65 ± 5% relative humidity. Maize grains of the variety “Plata” were used, and the grains were equilibrated to a moisture content of 12 to 13% at 30°C for insect rearing and use in the tests. All experiments were carried out in a dark room that was maintained at 30 ± 2°C and 65 ± 5% relative humidity.
Conditioned maize
The term “conditioned” is used here to refer to maize grains infested with stored-product insects (Mignon et al., 1996). Conditioned grains were obtained by placing approximately 150 unsexed S. zeamais adults in one-litre glass jars filled with 500 g of maize grains (approximately 1500 to 1600 grain kernels). After 14 to 21 days, the adults were removed and the maize was used for the experiments.
Egg plug identification
Sitophilus zeamais females deposit single eggs in holes bored into the grain, and seal each hole with a gelatinous egg plug. The egg plugs may be stained (cherry red) with acid fuchsin (Figure 1). The method used to stain the egg plugs was adapted from that of Holloway (1985). In brief, this method involves soaking grains for 1 to 2 min in warm water (25 to 30°C), followed by immersion for 1 to 2 min in an acid fuchsinic solution (0.5 g/L of water). The grains are finally rinsed with water to eliminate excess staining solution, and are then dried at room temperature. The egg plugs appear cherry red in colour when observed under a binocular microscope. We assumed that each plug covered just one egg and that the washing of kernels did not remove aggregation pheromone residues on infested kernels, if present.
Recording system
A Sterilin square box with 100 mm side exits was used (Bibby Sterilin Ltd, Stone, Staffs, UK). The box contains 25 square wells with 20 mm sides. One female of S. zeamais individuals from the laboratory cultures was placed in a well with three maize grains. The box was placed in a small dark room lit by a red light, not visible for insects (Chapman, 1998). In this room, a camera (Video-Flex, by Ken-a-vision, Raytown Road, Kansas City, Missouri, USA) connected to a video tape recorder (Panasonic, NV-SD430EG) was used to record insect displacement (Figure 2). Recordings took place between 08 and 09 am. After 6 h, the recording was stopped, and the displacement sequences were viewed on a colour monitor (JVC, TM1500PS). After each 5 min interval, we raised the grain on which the female is located.
Experimental procedures
Experiment 1: Progeny production in conditioned maize grains
This experiment was used to determine the number of progeny produced by the maize weevil on the two grain types (uninfested and conditioned). Seventeen male/female pairs were placed in 95 × 25 mm glass tubes containing 50 uninfested grains. A similar number of pairs were placed in identical tubes on the equivalent number of conditioned grains. Each tube was covered with a perforated plastic cap and kept in the dark room. The tubes were left undisturbed for 1 week, and then the adults were removed. The tubes were kept for 7 weeks, and then the emerged adults were sieved out and counted to provide a measure of productivity (number of emergent offspring). Productivity provides a reliable index of the number of eggs laid if there is no density-dependent mortality. This issue was not considered to be important in the present experiment; however, it was assessed because a large number of grains (50) were available for oviposition by each female over a relatively short period of 1 week. All of the grains used in this experiment were kept in a freezer at –20°C for 1 week to kill any larvae and other organisms that might have been present. The grains were then transferred to and maintained in the rearing chamber for 36 h to obtain water content equilibrium before being used for the experiment.
Experiment 2: Influence of conditioned maize on the selection of grains
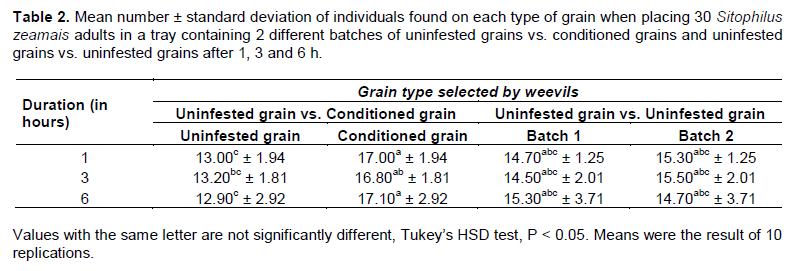
We placed 150 uninfected and 150 conditioned grains (42.2 to 45.3 g) side by side at a single-kernel thickness in a plastic tray (200 ×150 × 40 mm). Thirty unsexed S. zeamais adults were deposited at the centre of the tray. The tray was covered with a lid.
After 1, 3 and 6 h, the number of adults in each batch of grains was recorded (visually). Adults are mobile; thus, we assumed that 6 h is sufficient to judge grain preference. Ten replicates were made on 10 separate trays. The number of insects per grain type reflected the host selection preference of S. zeamais. The same procedure was repeated with two batches of uninfested grains to confirm that insect movement and distribution was not random.
Experiment 3: Selection of grain-hosts
The selection of grain-hosts by the maize weevil female was investigated under conditions containing one uninfested maize grain, one conditioned maize grain without egg and one conditioned maize grain of varying egg density (1, 2-3, 4 and 5 or more eggs/grain). The three different grains were exposed to an adult female weevil, which had been randomly selected from the laboratory cultures in a well. The insect displacement sequences were obtained by observing 6 h video recordings of the well, using the method described in Recording system. Eighteen to 24 replicates were performed for each egg density. The grain-host selection preference of S. zeamais was reflected in the number of times that the female was found on each type of grain.
Experiment 4: Effect of the presence of egg plugs on oviposition behaviour
Conditioned maize grains of varying egg density (0, 1, 2, 3 and 4 eggs/grain) were selected. At each density, five conditioned grains were mixed with five uninfested grains and placed on a Petri dish (55 mm Æ). The uninfested grains were stained with acid fuchsin to avoid bias, because the conditioned grains had been stained. Both grain types were marked using two different coloured dots made with an “Artline 725” permanent marker. Adult weevils were selected randomly from the laboratory culture, and the sexes were separated based on the dimorphic rostrum characteristics (Halstead, 1963) and were presumed fertile.
Ten S. zeamais adults (five females and five males) were added to the grains and the Petri dishes were placed in the dark room under controlled laboratory conditions (see Grain and Insects section). Nine replicates were performed at each egg density. After 3 days, the adults were removed and the grains were stained again with acid fuchsin. Then, the number of egg plugs was counted for the two grain types. The average number of eggs laid on the conditioned grains was obtained after subtracting the number of eggs found from the number of eggs initially present.
Statistical analysis
Data were subjected to analysis of variance (ANOVA), followed by Tukey’s HSD multiple comparison test at a = 0.05 when differences were detected in ANOVA, using the computer software SYSTAT. The square root and the angular transformation were used when variances were nonhomogeneous to correct for heterogeneity of variance prior to analysis (Dagnelie, 1986). However, the untransformed data are presented in the tables.
Progeny production on conditioned grains
The number of progeny produced was higher in previously conditioned maize. Significant differences (P = 0.004 < 0.05; F = 9.34; DF = 1) were observed in the means of adults present on conditioned and uninfested maize. The highest and lowest number of adults emerged on conditioned and uninfested maize (Table 1). Assuming that the number of emergent offspring represents a reliable index of eggs laid, this result implies that more eggs were laid on conditioned maize compared to
uninfested maize.
Influence of conditioned maize on S. zeamais host selection
Significantly, more S. zeamais adults were found on conditioned maize. The two-way ANOVA did not indicate any significant difference in the distribution of adult weevils at different observation times (P = 1 > 0.05; F = 0.001; DF = 3). In addition, a similar number of weevils were attracted to the two grain types in all three test periods. Adult weevil distributions differed on conditioned vs. uninfested grain (P = 0.0001 < 0.001; F = 13.35; DF = 2) (Table 2). More adults were present on conditioned grains. Adult distribution was similar when given the choice of 2 batches of uninfested grains (Table 2). Thus, we confirmed that insect movement was not random on conditioned vs. uninfested grains. Conditioned grains seemed to have an attractant effect on the maize weevil.
Selection of grain-hosts by S. zeamais
During the 6 h of recording, females were found at least 30 times on the conditioned grain carrying eggs, irrespective of the number of eggs. Females were also found on the other two grain types (uninfested and conditioned grain without eggs), averaging 12 to 18 times (Table 3). The two-way ANOVA (after the angular transformation of the data) showed significant differences (P = 0.0001; F = 28.65; DF = 2) between the two grain types. In contrast, there was no significant difference in the number of eggs carried by grains (P = 0.985; F = 0.01; DF = 2) and in the interaction of grain type-egg density (P = 0.816; F = 0.39; DF = 4). Thus, maize grains carrying eggs may attract female maize weevils.
Effect of the presence of egg plugs on S. zeamais oviposition behaviour
Females appeared to lay their eggs preferentially on conditioned grains (Table 3). The number of egg plugs on maize grains increased in relation to the number of previously deposited eggs. Gravid females oviposited more than 80% of eggs on grains containing two or more egg plugs. The two-way ANOVA showed significant differences (P = 0.0001; F = 26.543; DF = 1) between the two grain types and a significant difference among egg density levels (P = 0.031; F = 2.813; DF = 4) (Table 4). Conditioned grain may be preferred because of the density of eggs. The presence of a previously deposited egg may serve as a key attractant.
This study demonstrated that S. zeamais adults are able to differentiate between conditioned and uninfested maize during grain-host selection and egg-distribution on grain under laboratory conditions. Grains already conditioned by conspecific adults attracted subsequent maize weevils. More individuals emerged from conditioned maize because females laid more eggs on this grain type compared to uninfested maize. A similar phenomenon was observed by Longstaff (1981) which showed that S. zeamais clumps its eggs. This behaviour can be explained by the fact that egg aggregations occur because females prefer to oviposit on large grains. Yet, Stejskal and Kucerova (1996) showed that the oviposition preference for larger kernels is not constant, but is density dependent. For example, a decrease in egg aggregation occurs as the number of grains available for oviposition increases. In this study, grains were sorted, initially by both sieving and then visually, to avoid cracked grains, which attract maize weevils (Walgenbach et al., 1987), and to obtain uniformity in the shape and size of the grains. However, some conditioned kernels might have been slightly damaged or food odours may have attracted the adults.
Moreover, we found that S. zeamais females not only distinguish between conditioned and clean seeds, but also assess egg load quantitatively. Maize weevil females are able to recognise the presence of eggs and preferentially oviposit on grains with high egg density. The ability of females to perceive differences in egg density implies that relevant cues only exist after each preceding oviposition. A chemical basis of host recognition should be related to the aggregation pheromone (Walgenbach et al., 1983; Schmuff et al., 1984; Phillips et al., 1985; Walgenbach et al., 1987) called (4S,5R)-sitophinone (Walgenbach and Burkholder, 1986). Walgenbach et al. (1983) showed that when feeding on hosts, males produce an aggregation pheromone that attracts both sexes and causes "aggregation at optimum locations" where feeding, mating and oviposition might take place.
The aggregation pheromone also causes a larger number of individuals to gather locally, altering the microclimate, which would benefit the individual that released the pheromone (Plarre, 1998). Rising humidity and temperature have been recorded in aggregation hotspots formed by stored-product beetles and weevils (Longstaff, 1981; Sinha, 1984). It is likely that the conditioned grain in this experiment was contaminated with aggregation pheromone produced by male weevils. However, the deposition of eggs in the centre of an aggregation might endanger offspring, due to the presence of feeding adults. Mated females must strike a balance between favourable humidity and temperature conditions versus security for offspring, when selecting oviposition sites. According to Plarre (1998), the optimal location is on the periphery of hotspots. This phenomenon may explain why grain samples taken from the area surrounding a baited trap contained more weevil progeny than grain samples taken from the area surrounding unbaited traps (Plarre, 1996).
Preferred oviposition on grains already containing eggs was not expected, because studies with other insects that have confined larval stages (tephrids, bruchids, etc.) indicate that females should avoid competition and place eggs on grains that are uninfested (Messina and Renwick, 1985a, b). Previous works have shown that, as well as reducing the survival of larvae to maturity; larval competition also negatively influences the fecundity of surviving adults (Colegrave, 1993). In addition, larval competition does not influence adult fecundity, independent of its effect on adult weight (Credland et al., 1986; Colegrave, 1993). Consequently, our results raise two fundamental questions: how does this oviposition behaviour confer a fitness advantage; and how is it adaptive for the females that choose to cluster eggs in a stored grain environment.
If the tendency to clump eggs within grain does not depend on the experimental conditions, this type of oviposition strategy could be related to a certain adaptive mechanism. The presence of ovicide and cannibalistic behaviour represent two possible selective strategies. In general, weevil larvae are not able to migrate between grains and larval competition is assumed to be very high in a host grain containing multiple larvae. Either the Sitophilus female should (1) attempt to reduce individual competition faced by its offspring by killing eggs that are already present (ovicidal strategy), or (2) a larva within a grain should destroy other larvae (attack strategy) to maximise its expected fitness (Smith and Lessells, 1985). Regarding weevil behaviour, the hypothesis of the attack strategy is supported by the fact that larval cannibalism, which is known to enhance adult fecundity and generate nutritional benefit (Alabi et al., 2008; Alabi et al., 2009; Alabi, 2010; Via, 1999), seems to be controlled genetically (Stevens, 1989). However, there are no published estimates of ovicide and larvicide in Sitophilus species. Furthermore studies about cannibalism and fecundity may help provide explanations for the results reported in this paper.
The results of this study provide baseline information from which further research on cannibalism and fecundity of pest insect species inhabiting stored agricultural products may be investigated.
The author(s) have not declared any conflict of interest.
We thank the anonymous referees for their critical reviews during the preparation of this manuscript. This research was supported by a PhD grant from the Belgian Cooperation Agency to Danho Mathias.
REFERENCES
|
Alabi T (2010). Cannibalism and predation behaviours among Tribolium species (Coleoptera : Tenebrionidae). PhD thesis, Gembloux Agro-Bio Tech - University of Liege, Belgium. |
|
|
|
Alabi T, Michaud JP, Arnaud L, Haubruge E (2008). A comparative study of cannibalism and predation among seven Tribolium species (Coleopteran: Tenebrionidae). Ecol. Entomol. 33:716-726. |
|
|
|
Alabi T, Patigny S, Francis F, Haubruge E (2009). Origine et évolution du cannibalisme dans les populations animales: pourquoi manger son semblable. Biotechonol. Agron. Soc. Environ. 13:409-425. |
|
|
|
Chapman RF (1998). Visual signals : color and light production. In Chapman RF (eds) The insects structure and function. 4e edition, Cambridge University Press, U.K. |
|
|
Colegrave N (1993). Does larval competition affect fecundity independently of its effect on adult weight. Ecol. Entomol. 18:275-277.
CrossRef |
|
|
Credland PF, Dick KM, Wright AW (1986). Relationship between larval density, adult size and egg production in the cowpea seed beetle Callosobruchus maculatus. Ecol. Entomol. 11:41-50.
CrossRef |
|
|
Credland PF, Wright AW (1990). Oviposition deterrents of Callosobruchus maculatus (Coleoptera: Bruchidae). Physiol. Entomol. 15:285-298.
CrossRef |
|
|
|
Dagnelie P (1986). Théorie et méthodes statistiques. Applications agronomiques. Les presses agronomiques de Gembloux. Belgique. P. 2. |
|
|
|
Fava A, Burlando B (1995). Influence of female age and grain availability on the ovipositional pattern of the wheat weevil Sitophilus granarius (Coleoptera: Curculionidae). Eur. J. Entomol. 92:421-426. |
|
|
Fava A, Springhetti A (1991). Eggs distribution of Sitophilus granarius L. (Col., Curculionidae) in Triticum aestivum caryopses. J. Appl. Entomol. 111:406-411.
CrossRef |
|
|
Giga DP, Smith RH (1985). Oviposition markers in Callosobruchus maculatus (F.) and C. Rhodesianus (Pic.) (Coleoptera: Bruchidae): asymmetry of interspecific responses. Agric. Ecosyst. Environ. 12:229-233.
CrossRef |
|
|
Halstead DGH (1963). External sex differences in stored-products Coleoptera. Bull. Entomol. Res. 54:119-134.
CrossRef |
|
|
|
Holloway GJ (1984). Genetic differentiation and life history variation in the rice weevil, Sitophilus oryzae. PhD thesis, University of Reading, U.K. |
|
|
Holloway GJ (1985). The effect of increased grain moisture content on some life history characters of Sitophilus oryzae (L.) after staining egg plugs with acid fuchsin. J. Stored Prod. Res. 21:165-169.
CrossRef |
|
|
|
Longstaff BC (1981). Biology of the grain species of the genus Sitophilus (Coleoptera, Curculionidae): A critical review. Protect. Ecol. 2:83-130. |
|
|
MBata GN (1992). Egg distribution on seeds by Callosobruchus subinnotatus (Pic.) (Coleoptera: Bruchidae). J. Stored Prod. Res. 28:301-305.
CrossRef |
|
|
|
Messina FJ, Balmore JL, Renwick JAA (1987). Oviposition deterrent from eggs of Callosobruchus maculatus: spacing mechanism or artifact. J. Chem. Ecol. 13:219-226. http://dx.doi.org/10.1007/BF01020364PMid:24301372 |
|
|
Messina JF, Renwick JAA (1985a). Ability of ovipositing seed beetles to discriminate between seeds with differing egg loads. Ecol. Entomol. 10:225-230.
CrossRef |
|
|
Messina JF, Renwick JAA (1985b). Mechanism of recognition by the cowpea weevil Callosobruchus maculatus. Entomol. Exp. Appl. 37:241-245.
CrossRef |
|
|
Mignon J, Haubruge E, Liénad V, Gaspar C, Lognay G (1996). Mortality in Oryzaphilus surinamensis following short-term exposure to conditioned kernels by high-density culture. Entomol. Exp. Appl. 80:555-557.
CrossRef |
|
|
|
Nardon P, Grenier AM, Chessel D (1988). Comportement de ponte de la femelle de Sitophilus oryzae L. (Coléoptère: Curculionidé) : problème du choix des grains et du site de ponte sur le grain. Bull. Soc. Entomol. France 106:487-495. |
|
|
|
Phillips JK, Walgenbach CA, Klein JA, Burkholder WE, Schmuff NR, Fales HM (1985). (R*,S*)-5-hydroxy-4-methyl-3-heptanone: a male produced aggregation pheromone of Sitophilus oryzae (L.) and S. zeamais Motsch. J. Chem. Ecol. 11:1263-1274.
CrossRef
|
|
|
Plarre R (1996). Three dimensional distribution of Sitophilus granarius (L.) (Coleoptera, Curculionidae) in wheat influenced by the synthetic aggregation pheromone. J. Stored Prod. Res. 32:275-283.
CrossRef |
|
|
|
Plarre R (1998). Sex and aggregation pheromones of stored product insects. Different strategies with different biological background. Book of abstracts of the 2nd International Symposium on Insect Pheromones. WICC-International Agricultural Centre Wageningen, The Netherlands, 30 March-3 April, pp. 57-59. |
|
|
Schmuff NR, Phillips JK, Burkholder WE, Fales HM, Chen CW, Roller PP, Ma M (1984). The chemical identification of the rice weevil and maize weevil aggregation pheromone. Tetrahedon Lett. 25:1533-1534.
CrossRef |
|
|
Sinha RN (1984). Effects of weevil (Coleoptera, Curculionidae) infestation on abiotic and biotic quality of stored wheat. J. Econ. Entomol. 77:1483-1488.
CrossRef |
|
|
|
Smith RH (1986). Oviposition, competition and population dynamics in storage insects. Proceedings of the fourth International Working Conference on Stored-Product Protection, Tel Aviv, Israel, September, pp. 426-433. |
|
|
|
Smith RH, Lessells CM (1985). Oviposition, ovicide and larval competition in granivorous insects. Behavioural Ecology: ecological consequences of adaptative behaviour (ed. by R.M. Sibly and R.H. Smith), Blackwell Scientific Publications, Oxford, UK. pp. 423-448. |
|
|
Stejskal V, Kucerova Z (1996). The effect of grain size on the biology of Sitophilus granarius L. (Coleoptera : Curculionidae). I. Oviposition, distribution of eggs and adult emergence. J. Appl. Entomol. 120:143-146.
CrossRef |
|
|
Stevens L (1989). The genetics and evolution of cannibalism in flour beetles (genus Tribolium). Evolution 43:169-179.
CrossRef |
|
|
Via S (1999). Cannibalism facilitates the use of a novel environment in the flour beetle, Tribolium castaneum. Heredity 82:267-275.
CrossRef |
|
|
Walgenbach CA, Burkholder WE (1986). Factors affecting the response of the maize weevil, Sitophilus zeamais (Coleoptera, Curculionidae), to its aggregation pheromone. J. Environ. Entomol. 15:733-738.
CrossRef |
|
|
Walgenbach CA, Phillips JK, Faustini DL, Burkholder WE (1983). Male-produced aggregation pheromone of the maize weevil, Sitophilus zeamais and interspecific attraction between three Sitophilus species. J. Chem. Ecol. 9:831-841.
CrossRef |
|
|
Walgenbach CA, Burkholder WE, Curtis MJ, Khan ZH (1987). Laboratory trapping studies with Sitophilus zeamais (Coleoptera: Curculionidae). J. Econ. Entomol. 80:763-767.
CrossRef |