Full Length Research Paper

ABSTRACT
The effects of artificial biologically effective UV-B radiation on a range of growth and physiological parameters in two cotton (Gossypium hirsutum L.) cultivars (Romanos and Allegria) were recorded. Three levels of biologically effective UV-B were used: (1) zero (2) ambient and (3) elevated (determined as that associated with a notional 15% depletion of stratospheric ozone). Plants were grown under artificial light in growth chambers and subjected to the biologically effective UV-B radiation treatments. Compared to the zero level, the ambient and elevated biologically effective UV-B radiation significantly reduced plant height, leaf chlorophyll content, net photosynthetic rate, stomatal conductance, bract length, petal length, anthers number, pollen germination, seed cotton weight, fibre strength, fibre elongation, fibre micronaire, fibre maturity index, fibre spinning consistency index, mean fibre length, fibre yellowness and fibre uniformity index. Both the ambient and the elevated UV-B radiation also significantly increased stomatal density, short fibre index and fibre reflectance.
Key words: UV-B radiation, cotton, photosynthesis, stomatal density, flower characteristics, pollen germination, seed cotton weight, fibre quality.
INTRODUCTION
Chlorofluorocarbons (CFCs) released into the atmosphere in earlier years have depleted the stratospheric ozone layer resulting in increased levels of UV-B radiation reaching the Earth's surface (Rowland, 1990). As CFC levels gradually decline, the amount of UV-B radiation in the northern mid-latitudes is projected to return to its pre-1980 level by about 2065 (McKenzie et al., 2011).
Depending on location and season, crops grown between latitudes 40° N and 40° S currently receive a solar UV-B radiation dose of some 2-10 kJm-2 per day (Reddy et al., 2003). In several crops including cotton, a variety of plant processes are affected by UV-B radiation (Reddy et al., 2003; Kakani et al., 2003a). These processes include those associated with crop production, such as pollination, boll formation, boll development and lint yield.
Plants perceive UV-B by the specific ultraviolet-B photoreceptor UV Resistance Locus 8 (UVR8) that at the molecular level has been identified in Arabidopsis thaliana to enhance photomorphogenesis (Lee, 2016). It has also been reported that UVâ€B radiation affects crop yield and quality parameters (Wargent and Jordan, 2013).
In Greece, the cotton crop is an important component of trade and economy. However, the understanding of cotton’s response to elevated UV-B is limited, especially in relation to the plant’s growth and development, as well as in relation to fibre yield and quality (Gao et al., 2003).
The objectives of the present study were: i) to estimate the effects of UV-B radiation on cotton plant height, leaf chlorophyll content, net photosynthetic rate and stomatal conductance, stomatal density, stomatal length, stomatal width, bract length, petal length, staminal column length, anthers number, pollen germination and seed cotton weight in two commercial cultivars; and (ii) to quantify the effects of UV-B radiation on various key fibre properties.
MATERIALS AND METHODS
Two commercial cotton (Gossypium hirsutum L.) cultivars (cv., that is, Romanos and Αllegria) were used in the present study.
Growth chambers
The experiment was conducted in UV-B plant growth chambers at a controlled environment facility in University of Thessaly, Volos City, Greece. The chambers were lined internally with polyethylene sheets (TUV 3999, Crete plastics, Heraklion, Crete, Greece), which absorbed all UV-A and UV-B radiation. Sheets were replaced every two weeks. The chamber air conditioner comprised a cooling system, a heating system, a ventilation system and a control system. The minimum and maximum temperatures were 21-29°C during the light period and 15-19°C during the dark period. The relative humidity range was 41-65% (measured by a HOBO LCD data logger).
Each chamber was illuminated with multiple lamps emitting photosynthetic active radiation (PAR), UVâ€B radiation (280-315 nm) and ultraviolet A radiation (UV-A; 315-400 nm). The lamps were mounted at different heights above the plant canopy. Each chamber contained four PAR lights, being two of a metal halide type (MH; Osram HQI-TS 1000 W) and two of a high-pressure sodium type (HPS; Phillips SON-T 1000 W). Both lamp types were emitting some UV-A radiation. The average PAR measured using a 6200 quantum sensor (LI-COR, Lincoln, Nebraska, USA) just above the canopy ranged from 497.00 ± 90 μmol m-2 s-1 to 1042.00 ± 110 μmol m-2 s-1 during the experimental period (Table 1). The heights of the PAR lamps above the canopy were adjusted to the median plant height at least once per week to maintain a constant PAR exposure.
UV-B and PAR treatments
Three levels of biologically effective ultraviolet-B radiation (UV-BBE) per photoperiod duration were used: (1) 0 UV-BBE (control; 0 kj m-2day-1 UV-BBE) (2) Am UV-BBE (mean ambient UV-BBE radiation) and (3) UV-BBE 15 (mean enhanced UV-BBE radiation with a 15% reduction in stratospheric ozone). In the control chambers, the UV-B tubes were inactivated so that they did not emit UV-B radiation. The intensities of visible light and the degree of mutual shading from the UV tubes were similar across all chambers.
Solar UV-B and PAR values at ground level were determined for the city of Volos (Greece) using the MODerate resolution Imaging Spectroradiometer (MODIS) (http://modis.gsfc.nasa.gov/) onboard the satellites Terra and Aqua. The Radiative Transfer Model (LibRadtran) was used to simulate GHI and DNI for different scenarios of atmospheric parameters. Spectra of solar UV-B irradiance reaching the ground were calculated. These spectra were weighted with the Caldwell (1971) generalised plant action spectrum (normalised at 300 nm) to determine the biologically effective UV-B radiation dose. The average values of photoperiod duration and photosynthetically active radiation (PAR) were also calculated.
Throughout the period from seedling emergence to boll harvest, the mean Am UV-BBE and UV- BBE15 were changed according to the duration of photoperiods. Also, the mean PAR values during the photoperiod and the mean photoperiod length were changed (Table 1). The mean UV-A radiation values were supplemented by four Phillips TLD 36W/08 lamps, which together with the metal halide and high-pressure sodium lamps provided a total UV-A flux of 1.27 ± 0.05 W m-2 at plant height. The UV-BBE radiation was delivered to the plants during the photoperiod; the doses are listed in Table 1.

In each chamber, the heights of the PAR, UV-B and UV-A lamps were adjusted to median plant height once per week. Two types of polyethylene sheets (Crete plastics, Heraklion, Crete, Greece) were used: a) TUV 3942 filter that blocks UV-B and transmits UV-A and longer wavelength radiation in control chambers and b) TUV 3999 filter. The UV-BBE radiation was provided by five parallel fluorescent tubes (TL 40 W/12 RS-Philips, Holland) perpendicular to the plant rows and 0.6 m above the canopy. The energy of the emitted UV-B and UV-A radiations were checked, adjusted and delivered according to photoperiod durations, using a computer, UV-B and UV-A sensors and microcontrollers. The fluorescent UV-B light was filtered through 75 μm thick cellulose diacetate sheets (Clarifoil, Coventry, UK). Cellulose diacetate was used to eliminate any UV-C radiation but to transmit UV-BBE and longer wavelengths, including UV-A radiation. Cellulose diacetate filters were replaced every 2 days to ensure uniformity of UV-B and UV-A transmission due to photodegradatiοn. UV-B and UV-A irradiances were measured using SKU 430 and SKU-420 sensors (Skye Instruments, Llandrindod Wells, UK), respectively.
Plant culture and growth conditions
The experiment began one day after emergence (1 DAE) and continued to harvest (172 DAE) (Table 1). Cotton seeds of the two cultivars were sown in a substrate of 80:20 (w/w) soil:peat in rectangular steel bins on wheels, equipped with drain holes 810 (high) x 770 (long) x 230 mm (wide). The main characteristics of the soil were: colour 10YR 3/3, sand 31%, silt 34%, clay 35%, texture Clay Loam, organic matter 1.77 g/100 g soil, CaCO3 10.1%, pH 7.8 (H2O 1:1), phosphate 10 ppm (Olsen). The exchangeable cations were: K 0.45, Na 0.17, Ca 30.15 and Mg 7.83 me/100 g soil (Mitsios et al., 2000). The peat used was Floradur R: pH 5.0-6.5, N 50-300, P2O5 80-300 and K2O 80-400 mg/l) produced by Floragard Vertriebs GmbH für Gartenbau. After germination, seedlings were thinned to four plants per pot, with main stems about 170 mm apart. Plants received about 3 L of tap water per day via a dripper. A complete fertiliser (Hakaphos N:P:K, 12:32:14+3% Mg, Compo Hellas) was added on three occasions on 27, 57 and 99 DAE. To minimize positional effects, pots were completely randomized within each chamber every third day.
Measurements
Plant height was determined on eight plants per UV-BBE treatment and cultivar on 92 and 170 days after emergence. Total chlorophyll contents were estimated in the same leaves after net photosynthetic rates and stomatal conductance determination. Contents of total chlorophyll (chlorophyll a and chlorophyll b) of fully-expanded leaves from the top of the main stem were determined in SPAD units by a chlorophyll meter (SPAD 502, Minolta LTD, Ojaka, Japan). Eight leaves, derived from 8 plants by different pots, were used per UV-B treatment and cultivar. Eight average values were taken from each leaf.
Net photosynthetic rate (Pn) and stomatal conductance (gs) were measured per UV-B treatment and cultivar on the youngest, fully expanded mainstem leaves from the top of the main stem of eight plants, from different pots. These measurements were made using a LI 6200 portable photosynthesis system (LI-COR, Lincoln, NE USA).
Portions of the adaxial and abaxial surfaces of the youngest, fully expanded mainstem leaves (counting from the top of the main stem) were coated with clear nail varnish, in the mid-area between the central vein and the edge of the leaf. The chosen leaves were those for which gas exchange and chlorophyll content were measured. After drying, the peels were removed using fine forceps and placed on a slide. The numbers of stomata per mm2 (stomatal density), stomatal length and stomatal width were counted using a light microscope (x400 magnification). The numbers of stomata were counted in three fields on each leaf surface of eight replicate leaves per treatment and cultivar. The leaves were selected from eight plants from different pots. Stomatal length and stomatal width of randomly chosen stomata was also measured in three different fields on each leaf surface of eight leaf samples from eight plants, under the same magnification using an ocular micrometer. Therefore, the length or width of 24 stomata for each UV-B treatment, cultivar and leaf surface was measured.
Total chlorophyll content, photosynthetic, stomatal conductance, stomatal density, stomatal length and stomatal width were measured on 66 and 128 days after emergence.
Lengths of bracts, petals, maximum measurable length of staminal columns and anther number for five flowers in each treatment and cultivar were counted on 71 and 91 days after emergence
Early in the morning on 67 and 88 days after emergence, when the anthers were beginning to dehisce and pollen grains were at the same developmental stage, five flowers were randomly selected from five plants per treatment and cultivar by plants grown outdoors. Flower peduncles were immediately placed with their cut ends in a porous material (Oasis floral foam blocks) impregnated with tap water. The flowers were placed under lamps of 0 UV-BBE, Am UV-BBE, and UV-BBE 15 radiation at 0.60 m. Then, 5-6 anthers were taken from each flower and transferred to a drop of liquid germination medium on a glass slide. This medium was made up according to Brewbaker and Kwack (1963) and modified to contain H3BO3 0.1 gl-1, Ca (NO3)2.4H2O 0.3 gl-1, Mg SO4 0.2 gl-1, KNO3 0.1 gl-1, KH2PO4 0.1 gl-1 and sucrose 100 gl-¹. Pollen grains were distributed uniformly in the drop of liquid medium using a needle. Slides were kept placed in moist Petri dishes, covered and incubated at 22±2°C for 2 h before storage at 4°C pending observation of germination by light microscopy. Pollen grains were considered to have germinated artificially when the pollen tube length was at least equal to or greater than one pollen grain diameter. Six microscope fields were examined per flower, cultivar and UV-BBE level, with each field containing 30-70 pollen grains. Germination percentage was determined as the fraction of the total grains present.
Seed cotton weight per plant was determined at 171 days after emergence. Samples of seed cotton from eight plants were gathered by hand and put in plastic bags. Qualitative characteristics of cotton fibres were determined in the high volume instrument machine in the laboratory of a local textile industry, after harvesting the raw cotton from eight plants.
Statistical analyses
The collected data was computerized and analyzed using ANOVA to determine least significant differences (LSD tests). The software package IBM SPSS Statistics V23.0 was used for all statistical computations.
RESULTS AND DISCUSSION
The UV-B treatments significantly affected all traits of both cultivars (Tables 2 to 6). The pollen germination and fibre quality results reported in the present study are the first describing ambient and supplemental UV-BBE radiation effects on cotton.




Plant height
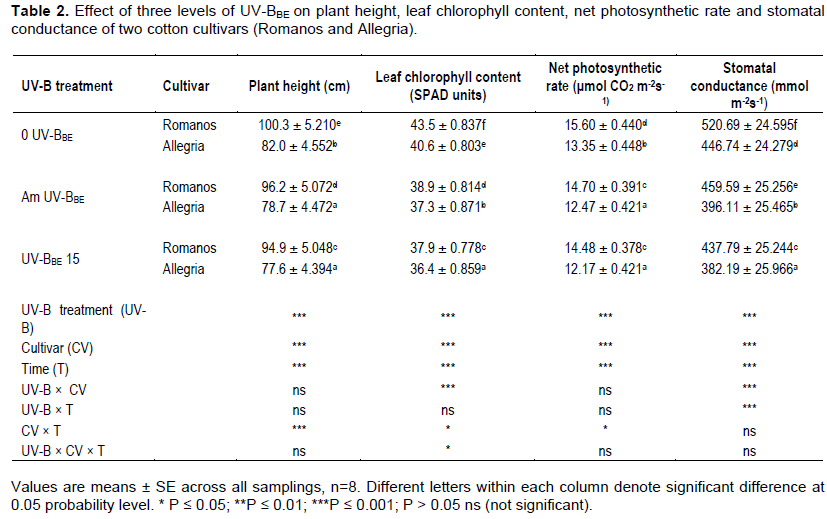
Throughout the experimental period, plant height was reduced by increased UV-BBE irradiance. In both cultivars, decreases in plant height due to UV-BBE followed the pattern: plant height under 0 UV-BBE > Am UV-BBE > UV-BBE 15. Throughout the sampling period, compared with the controls, the plant height reductions in cv. Romanos were 4.09% (Am UV-BBE) and 5.38% (UV-BBE15) and in cv. Allegria were 4.02% (Am UV-BBE) and 5.37% (UV-BBE15) (Table 2). These results are consistent with those of Gao et al. (2003), who reported reductions in cotton plant height under field conditions due to enhanced UV-B radiation. The exclusion of solar UV-B radiation increased the specific leaf weight compared to the control and increased plant height with a significant biomass increase (Zhu and Yang, 2015).
In addition, UV-B treatments decreased plant height. The reduction in plant height following exposure to UV-B radiation has been attributed to inhibition of biosynthesis or signaling of various hormones (Vanhaelewyn et al., 2016), to reductions in indole-3-acetic acid (IAA) by increasing peroxidase and IAA oxidase activities that can cause growth suppression (Huang et al., 1997). The reductions are due to a shortening of internode length, rather to fewer internodes (Reddy et al., 2003).
Chlorophyll (a+b) content
In both cultivars, the mean chlorophyll content under exposure to the three UV-BBE levels followed the following reduced range: chlorophyll content of 0 UV-BBE > chlorophyll content of Am UV-BBE > chlorophyll content of UV-BBE 15. Leaf chlorophyll content was higher in cv. Romanos than in cv. Allegria in all UV-BBE equivalent regimes. Compared to controls, the leaf chlorophyll content reductions in cv. Romanos were 10.57% (Am UV-BBE) and 12.87% (UV-BBE 15) and in cv. Allegria were 8.13% (Am UV-BBE) and 10.34% (UV-BBE 15) (Table 2).
Kakani et al. (2003b) reported that chlorophyll content reduction after exposure to UV-B ranged from 10 to 78% among the dicot species. Enhanced UVâ€B radiation exposure significantly reduced total chlorophyll content, depending on crop cultivars/species, treatment doses, length and intensity of UV-B radiation, and variation in the PAR/UV-B ratio. The decrease in chlorophyll content was due to thylakoids and grana rupture on UV-B radiation exposure. Also, the declines in pigments of chlorophyll and photosynthesis led in reduced biomass and yield for most crop plants. Enhanced UV-B radiation due to depletion of stratospheric O3 decreases the financial yield and product quality of field crops.
Ambient UV-B radiation resulted in increased UV-B absorbing leaf compounds, while chlorophyll a, b, and (a+b) content reduced. However, chlorophyll bleaching and damage by solar UV-B radiation to the photosynthetic apparatus induced a decrease in photosynthetic rate (Zhu and Yang, 2015). It has been reported that chl-a and chl-b concentration in plants exposed to UV radiation has been significantly reduced. This is due to enhanced chlorophyll photodegradation, to lower chlorophyll synthesis rates resulting from decreased expression of genes encoding chlorophyll-binding proteins, or to break down of chloroplasts structural integrity (Sarghein et al., 2008). The previously mentioned are the possible reasons for the reduction in the present study
Net photosynthetic rate
Compared to controls plants, the reduction of Pn under Am UV-BBE exposure was during the sampling 5.77% in Romanοs and 6.59% in Allegria, while under UV-BBE15 exposure the reductions were 7.18% for cv. Romanos and 8.84% for cv. Allegria. In both cultivars, net photosynthetic rate reduced under Am UV-BBE and even more under UV-BBE15. The variation of Pn following exposure to UV-B radiation in both cultivars usually followed the following pattern throughout the sampling period: Pn under 0 UV-BBE > Pn under Am UV-BBE > Pn under enhanced UV-BBE15 (Table 2).
It is inferred that the photosynthesis decline was closely correlated with a decline in stomatal conductance (Kakani et al., 2004). It was also shown that increased UV-B significantly decreased net photosynthetic rate and stomatal conductance (Yao and Liu, 2006). UV-B radiation exclusion significantly increased the net photosynthetic rate, stomatal conductance and activity of Rubisco. There was also a suppressive action of ambient UV-B on growth and photosynthesis; and dicots (as cotton) were more susceptible than monocots in this suppression (Kataria et al., 2013).
The effect of UV-B radiation on photosynthesis has been shown to depend on species of crops, cultivars, experimental conditions, dosage of UV-B, ratio of PAR to UV-B radiation, stages of plant growth and interactions between UV-B radiation and other stresses of the environment. The decrease in photosynthesis by UV-B doses ranged from 3-90% in crop plants. Decrease in photosynthesis was due to impact of UV-B on Photosystem II along with reduction in pigments and leaf area. However, the declines in pigments of chlorophyll and photosynthesis led in reduced biomass and yield for most crop plants (Kakani et al., 2003b).
Photosynthesis, however, can also be depleted by stomatal density and opening, reduced stomatal conductance or reduced chlorophyll content, following exposure of certain plants to UV radiation (Salama et al., 2011). Most of the rice cultivars plants grown in the greenhouse, exposed to enhanced UV-B radiation showed reduced photosynthetic rate, pollen germination, fertility and yield (Mohammed and Tarpley, 2011). UV-B radiation causes harm to the photosynthetic apparatus of green plants at various sites. UVB radiation has been shown to cause a decrease in photosynthetic activity primarily associated with the PSII protein degradation, chlorophyll and carotenoid destruction, reduced stomatal function activity and impacts on Rubisco activity (Kataria et al., 2014).
Stomatal conductance
During the sampling period, stomatal conductance was higher in cv. Romanos at similar UV-BBE intensities than in cv. Allegria. Also, gs under exposure to UV-B levels followed in both cultivars the following reduced scale: gs under 0 UV-BBE > gs under Am UV-BBE > gs under enhanced UV-BBE15. Compared to control plants, the reduction of stomatal conductance under Am UV-BBE exposure was during the sampling 11.73% in Romanοs and 11.33% in Allegria, while under UV-BBE15 exposure, the reductions were 15.92% for cv. Romanos and 14.45% for cv. Allegria. Plants under 0 UV - BBE showed a significant increase in the net rate of photosynthesis and stomatal conductance (Table 2). The results of Kataria et al. (2013) were similar after exclusion of the UV-B.
It has been reported that UV-induced changes in stomatal conductance reduced CO2 assimilation, because it affects the opening or closing of stomata through alterations in the stomatal aperture. It has been postulated that high fluences of UVâ€B either stimulated stomatal opening or stomatal closing in Vicia faba, depending on the metabolic state of the guard cell, and neither of these responses is readily reversed. High-UV-B acted on the guard cell aperture control and changed the mesophyll photosynthesis. High-UV-damaged the PSII in the guard cells, affecting photophosphorylation and hence ion transport, controlled osmotic solute flux, notably K+, from guard cells and the resultant changes in guard cell turgor and stomatal aperture. The plasmalemma based enzyme complexes facilitate the solute fluxes leading to stomatal opening and closure (Jansen and Van Den Noort, 2000). It is remarkable that UVR8 controls the stomatal closure by means of a mechanism involving both H2O2 and NO generation, which increased in UV-B-irradiated stomata, although stomatal closure required only NO (Tossi et al., 2014).
Stomatal density
Stomatal density was always higher on abaxial than on adaxial leaf surface of two cultivars. In both cultivars, both Am UV-BBE and enhanced UV-BBE 15 radiation increased the number of stomata on the adaxial or abaxial surfaces compared with the controls. The exposure of plants to UV-BBE radiation in both cultivars increased the stomatal density, with values to follow the following pattern across radiation levels: stomatal density under UV-BBE 15 > stomatal density under Am UV-BBE > stomatal density 0 UV-BBE. Compared with the zero UV-B controls, high UV-B treatment increased the number of stomata on the adaxial surfaces of cv. Romanos by 32.52% and of cv. Allegria by 23.74%. The corresponding increases on the abaxial surfaces were 13.00% for cv. Romanos and 11.96% for cv. Allegria (Table 3).
It has been reported that the UV-B enhancement included changes in the stomatal density, leaf area, leaf thickness, wax deposition, elongation of the stem and pattern of branching, in plant–pathogen and plant–predator interactions and gene expression as well as in the synthesis of secondary metabolites (Prado et al., 2012). In the present study, the increased stomatal density by UV-B radiation may have provided cotton cultivars with greater CO2 concentrations, which will increase their photosynthetic rates. Leaf area and net photosynthetic rates in cotton were reduced by enhanced UV-B radiation (Zhao et al., 2004). Thus, the distribution of the number of stomata over a smaller leaf area surface in both cultivars, increased stomatal density. Indeed, it has been reported that the stomatal density and conductance affect the CO2 uptake and, therefore, photosynthesis (Zheng and Van Labeke, 2017).
Stomatal length and width
There were no significant differences in stomatal length and stomatal width among UV-B levels (Table 3).
Flower characteristics
Bract length, petal length and anther number were significantly reduced by the ambient UVâ€BBE and UV-B BE15 treatments compared with greatest values for plants in the controls (Table 4). Throughout the sampling period, compared with the controls, the anther reductions in cv. Romanos were 9.36% (Am UV-BBE) and 12.26% (UV-BBE 15) and in cv. Allegria were 7.98% (Am UV-BBE) and 10.93% (UV-BBE15), respectively (Table 4). There were also no significant differences between the flowers exposed to ambient UVâ€BBE and UV-BBE15 radiation treatments in the staminal column length (Table 4).
Pollen germination
Compared to the controls, both Am UV-BBE and enhanced UV-BBE 15 radiation reduced the mean pollen germination over cultivars sampling. With respect to cultivars, pollen germination in cv. Allegria was on average higher than in cv. Romanos. The data showed significant reductions in pollen germination for both genotypes. The pollen germination reductions in cv. Romanos were 32.61% (Am UV-BBE) and 45.68% (UV-BBE 15) and in cv. Allegria were 32.85% (Am UV-BBE) and 41.22% (UV-BBE 15) (Table 4).
According to Llorens et al. (2015), pollen grains are shielded by bracts or petals in entomophilous plants. As a consequence of having constitutively greater levels of UV-B protective compounds, ovaries are better shielded against UV-B radiation than other floral parts. Pollen is the reproductive tissue most susceptible to UV-B, particularly during anther dehiscence and pollen tube penetration of the stigma. There is also a tendency in annual species to reduce fruit and/or seed production as UV-B doses increase.
Increased UVâ€B radiation reduced in vitro the rate of pollen germination and tube length as well as its ability to fertilize in the field. Oxygen species (O2•− and H2O2) production increased with UVâ€B radiation and their ongoing accumulation resulted in lipid peroxidation and reduced antioxidant activity in maize (Wang et al., 2010).
He et al. (2007) observed that Paulownia tomentosa pollen exposed in vitro to UV-B radiation reduced pollen germination and tube growth, but also increased NO synthase activity and NO production in pollen grain and tube. UV-B radiation in maize pollen grains induced a significant increase in UV-B absorbing pigments (plants adaptation to complete their reproductive cycle) (Santos et al., 1998).
Seed cotton weight
Seed cotton weight was reduced by the UV-B, but the magnitude of the response was not similar across cultivars. Plant exposure to UV-B radiation resulted in reduced seed cotton weight compared with the controls, an effect that was more evident under the enhanced UV-BBE15 level.
Compared to the 0 UV-BBE level, the reductions in seed cotton weight under Am UV-BBE exposure were 2.91% in cv. Romanos and 4.24% in cv. Allegria, while the decreases were 3.45% in cv. Romanos and 5.27% in cv.
Allegria under the enhanced UV-BBE15 exposure (Table 5). Similarly, the supplemental UV-B irradiance declined significantly the unginned cotton yield (Gao et al., 2003). In addition, the exposure of plants to UV-BBE radiation resulted in lower seed weight and the magnitude of the reduction was dependent on UV-BBE level.
Lint quality traits
In comparison with the controls, fibre qualitative characteristics were reduced or increased by exposure to UV-B. The two cultivars also performed differently. Maximum reductions under UV-BBE15 were observed on fibre strength 10.48% (Romanos), elongation 23.49% (Romanos), micronaire 9.57% (Romanos), maturity index 4.65% (Romanos), spinning consistency index 4.03% (Romanos), mean length 2.71% (Allegria), yellowness 11.87% (Romanos) and uniformity index 3.8% (Romanos). Also, compared with the controls, maximum increases under UV-BBE15 were observed on fibre short index 18.01% (Romanos) and reflectance 10.3% (Allegria) (Tables 5 and 6).
Gao et al. (2003) found similar negative effects on cotton fibre quality under enhanced UV-B. Exposure to ambient UV-B radiation reduces the crops photosynthesis, growth, production of dry matter, yield and quality of grain (Gao et al., 2010). Changes in the yield and quality of wheat induced by increased UV-B throughout the whole growth stage (Yao et al., 2014). In addition, due to elevated UV-B, the seed quality of soybean cultivars was deteriorated (Choudhary and Agrawal, 2015). Under increased UV-B radiation, the protein content of maize grains was increased, but the content of oil and starch were not influenced (Yin and Wang, 2012). Furthermore, the most important flavor compounds of holy basil (Ocimum sanctum L) plants cultivated in the field after the biologically effective supplemental ultraviolet-B radiation treatment significantly increased (Kumari and Agrawal, 2011).
CONCLUSIONS
Both ambient UV-BBE and enhanced UV-BBE 15 irradiances significantly affected most of the cotton growth, physiological and fibre quality traits measured in the present study, with the higher UV-B dose generally having the strongest effect. Compared to the control, plants exposed to biologically effective UV-B radiation showed lower values in most traits, including pollen germination and fibre elongation, micronaire, spinning consistency index, and uniformity index. On the contrary, the values of stomatal density, short fibre index and fibre reflectance were increased due to the ambient and enhanced UV-B radiation compared to the control. There have been differences in cultivar response to UV-BBE in several cases, suggesting differential genotypic sensitivity of cotton to increased levels of UV-B radiation.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Brewbaker JL, Kwack BH (1963). The essential role of calcium ion in pollen germination and pollen tube growth. American Journal of Botany 50(9):859-865. |
|
|
Caldwell MM (1971). Solar UV irradiation and the growth and development of higher plants. Photophysiology 6:131-177. |
|
|
Choudhary KK, Agrawal SB (2015). Effect of elevated ultraviolet-B on four tropical soybean cultivars: quantitative and qualitative aspects with special emphasis on gas exchange, chlorophyll fluorescence, biomass and yield. Acta Physiologiae Plantarum 37(2):31. |
|
|
Gao W, Schmoldt DL, Slusser JR (2010). UV Radiation in Global Climate Change, Measurements, Modelling and Effects on Ecosystems. Tsinghua University Press, Beijing. |
|
|
Gao W, Zheng Y, Slusser JR, Heisler GM (2003). Impact of enhanced ultraviolet-B irradiance on cotton growth, development, yield, and qualities under field conditions. Agricultural and Forest Meteorology 120(1-4):241-248. |
|
|
He JM, Bai XL, Wang RB, Cao B, She XP (2007). The involvement of nitric oxide in ultravioletâ€bâ€inhibited pollen germination and tube growth of Paulownia tomentosa in vitro. Physiologia Plantarum 131(2):273-282. |
|
|
Huang S, Dai Q, Peng S, Chavez AQ, Miranda ML, Visperas RM, Vergara BS (1997). Influence of supplemental ultraviolet-B on indoleacetic acid and calmodulin in the leaves of rice (Oryza sativa L.). Plant Growth Regulation, 21(1):59-64. |
|
|
Jansen MA, Van Den Noort RE (2000). Ultravioletâ€B radiation induces complex alterations in stomatal behaviour. Physiologia Plantarum 110(2):189-94. |
|
|
Kakani VG, Reddy KR, Zhao D, Gao W (2004). Senescence and hyperspectral reflectance of cotton leaves exposed to ultravioletâ€B radiation and carbon dioxide. Physiologia Plantarum 121(2):250-257. |
|
|
Kakani VG, Reddy KR, Zhao D, Mohammed AR (2003a). Effects of ultravioletâ€B radiation on cotton (Gossypium hirsutum L.) morphology and anatomy. Annals of Botany 91(7):817-826. |
|
|
Kakani VG, Reddy KR, Zhao D, Sailaja K (2003b). Field crop responses to ultraviolet-B radiation: a review. Agricultural and Forest Meteorology 120(1-4):191-218. |
|
|
Kataria S, Guruprasad KN, Ahuja S, Singh B (2013). Enhancement of growth, photosynthetic performance and yield by exclusion of ambient UV components in C3 and C4 plants. Journal of Photochemistry and Photobiology B: Biology 127:140-152. |
|
|
Kataria S, Jajoo A, Guruprasad KN (2014). Impact of increasing Ultraviolet-B (UV-B) radiation on photosynthetic processes. Journal of Photochemistry and Photobiology B: Biology 137:55-66. |
|
|
Kumari R, Agrawal SB (2011). Comparative analysis of essential oil composition and oil containing glands in Ocimum sanctum L. (Holy basil) under ambient and supplemental level of UV-B through gas chromatography-mass spectrometry and scanning electron microscopy. Acta Physiologiae Plantarum 33(4):1093-1101 |
|
|
Lee JH (2016). UV-B signal transduction pathway in Arabidopsis. Journal of Plant Biology 59(3):223-230. |
|
|
Llorens L, Badenes-Pérez FR, Julkunen-Tiitto R, Zidorn C, Fereres A, Jansen MA (2015). The role of UV-B radiation in plant sexual reproduction. Perspectives in Plant Ecology, Evolution and Systematics 17(3):243-254. |
|
|
McKenzie RL, Aucamp PJ, Bais AF, Björn LO, Ilyas M, Madronich S (2011). Ozone depletion and climate change: impacts on UV radiation. Photochemical and Photobiological Sciences 10(2):182-198. |
|
|
Mitsios IK, Toulios L, Charoulis A, Gatsios FA, Floras SA (2000). Soil survey and soil map of Thessaly University farm, in Velestinon area. Zymel editions, Athens. (In Greek). |
|
|
Mohammed AR, Tarpley L (2011). Morphological and physiological responses of nine southern US rice cultivars differing in their tolerance to enhanced ultraviolet-B radiation. Environmental and Experimental Botany 70(2-3):174-84. |
|
|
Prado FE, Rosa M, Prado C, Podazza G, Interdonato R, González JA, Hilal M (2012). UV-B Radiation, its Effects and Defense Mechanisms in Terrestrial Plants. In. Parvaiz A, Prasad MNV (Eds.), Environmental Adaptations and Stress Tolerance of Plants in the Era of Climate Change. New York, NY: Springer Science Business Media pp. 57-83. |
|
|
Reddy KR, Kakani VG, Zhao D, Mohammed AR, Gao W (2003). Cotton responses to ultraviolet-B radiation: experimentation and algorithm development. Agricultural and Forest Meteorology 120(1-4):249-65. |
|
|
Rowland FS (1990). Stratospheric ozone depletion by chlorofluorocarbons. Ambio 19(6-7):281-292. |
|
|
Salama HM, Al Watban AA, Al-Fughom AT (2011). Effect of ultraviolet radiation on chlorophyll, carotenoid, protein and proline contents of some annual desert plants. Saudi Journal of Biological Sciences 18(1):79-86. |
|
|
Santos A, Almeida JM, Santos I, Salema R (1998). Biochemical and ultrastructural changes in pollen of Zea mays L. grown under enhanced UV-B radiation. Annals of Botany 82(5):641-645. |
|
|
Sarghein SH, Carapetian J, Khara J (2008). Effects of UV-radiation on photosynthetic pigments and UV absorbing compounds in Capsicum longum (L.). International Journal of Botany, pp. 486-490. |
|
|
Tossi V, Lamattina L, Jenkins GI, Cassia RO (2014). Ultraviolet-B-induced stomatal closure in Arabidopsis is regulated by the UV resistance locus8 photoreceptor in a nitric oxide-dependent mechanism. Plant Physiology 164(4):2220-2230. |
|
|
Vanhaelewyn L, Prinsen E, Van Der Straeten D, Vandenbussche F (2016). Hormone-controlled UV-B responses in plants. Journal of Experimental Botany 67(15):4469-4482. |
|
|
Wang S, Xie B, Yin L, Duan L, Li Z, Egrinya Eneji A, Tsuji W, Tsunekawa A (2010). Increased UVâ€B radiation affects the viability, reactive oxygen species accumulation and antioxidant enzyme activities in maize (Zea mays L.) pollen. Photochemistry and Photobiology 86(1):110-116. |
|
|
Wargent JJ, Jordan BR (2013). From ozone depletion to agriculture: understanding the role of UV radiation in sustainable crop production. New Phytologist 197(4):1058-1076. |
|
|
Yao X, Chu J, He X, Si C (2014). Grain yield, starch, protein, and nutritional element concentrations of winter wheat exposed to enhanced UV-B during different growth stages. Journal of Cereal Science 60(1):31-36. |
|
|
Yao X, Liu Q (2006). Changes in morphological, photosynthetic and physiological responses of Mono Maple seedlings to enhanced UV-B and to nitrogen addition. Plant Growth Regulation 50(2-3):165-177. |
|
|
Yin LN, Wang SW (2012). Modulated increased UV-B radiation affects crop growth and grain yield and quality of maize in the field. Photosynthetica 50(4):595-601. |
|
|
Zhao D, Reddy KR, Kakani VG, Mohammed AR, Read JJ, Gao W (2004). Leaf and canopy photosynthetic characteristics of cotton (Gossypium hirsutum) under elevated CO2 concentration and UV-B radiation. Journal of Plant Physiology 161(5):581-590. |
|
|
Zheng L, Van Labeke MC (2017). Long-term effects of red-and blue-light emitting diodes on leaf anatomy and photosynthetic efficiency of three ornamental pot plants. Frontiers in Plant Science 8:1-12. |
|
|
Zhu PJ, Yang L (2015). Ambient UV-B radiation inhibits the growth and physiology of Brassica napus L. on the Qinghai-Tibetan plateau. Field Crops Research 171:79-85. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


