Full Length Research Paper

ABSTRACT
The purpose of this study was to investigate the morphological diversity of plantain accessions from three main plantain growing districts in Benin Republic. Sixteen descriptors were used including 5 quantitative. Overall, 31 cultivars were identified and associated with various names depending on the sociolinguistic groups. Results indicated the presence in each district of the three types of inflorescences (male rachis appearance) that describe the plantain subgroup. False horn is the most represented with 19 accessions, followed by French type with 10 accessions and True horn with only two accessions. All plantains were giant cultivars with pseudostem greater than 3 m, except hybrid L5449, which averaged 2.89±0.06 m. The collection was dominated by accessions having slightly oblique or sub-horizontal bunches (83.87%). Quantitative variables were submitted to Principal Component Analysis (PCA) which led to the separation of all the accessions into four clusters having specific traits that can be harnessed diversely in a breeding program. The presence of the three types of inflorescences confirmed plantain phenotypical diversity, with “False horn” type dominating plantain cultivars. Moreover, the occurrence of genetic erosion of white pulp cultivars is due to consumer preferences.
Key words: Accession, collection of bananas, biodiversity, phenotypical variability.
INTRODUCTION
Plantain (Musa species AAB) is an essential component of the diet of populations in West and Central Africa, which is used to address food security challenge (Folefack et al., 2017). Plantain production was estimated annually at 11 million tons, representing more than 70% of total banana production in those areas (Lescot, 2020). This region has been recognised to host the highest plantain variability in the world with more than 150 cultivars. As such, West and Central Africa is considered as a second pole of plantain diversification, although Musa spp. are native to Southeast Asia (Blomme et al., 2013; Lescot 2010). Apart from the high diversity of local cultivars, attempts to solve pest problems related to plantain production have led to developing new hybrid varieties through breeding programs (Dochez et al., 2013; Kimunye et al., 2021; Tenkouano et al., 2019), while increasing the diversity of plantain varieties (Ortiz and Swennen, 2014). In this regard, data recorded on some cultivars cannot automatically be generalized to all available plantains, due to the lack of the right taxonomic and agro-morphological description of the wide diversity of plantains (Adheka et al., 2018). Furthermore, local names of those cultivars differ from one community to another, preventing research findings to be used by some communities that are not accustomed to those names (Saraswathi et al., 2011). This is a common reality in Benin where Musa is mainly grown by small-scale farmers in the southern and central regions of the country (Chabi et al., 2018). Most plantations in this country are extensive with no special care (Lokossou et al., 2012). Those plantations constitute a pool from which planting materials are selected to establishing new plantations (Ademiluyi, 2013; Sivirihauma et al., 2017a). In such a context, cultivars of different size coming from various fields are mixed together in the same plantation (Dépigny et al., 2016). Furthermore, production system are undermined by several constraints including the lack of knowledge with regard to planting activity and cares during growth and development stages, as well as to the selection of plantain cultivars that suit the context (Odah et al., 2013).
Currently, cultivar selection is mainly guided by farmers' behaviours and endogenous knowledge rather than standardized knowledge based on their performance (Dépigny et al., 2016). In this context, building farmer capacity with cultivar-specific knowledge would be a first step toward improving production of cropping systems, which requires the description and classification of germplasm based on agro-morphological characterization (Mohamed et al., 2019). In fact, morphological traits have been used over the past 70 years in the study of Musa diversity (Adheka et al., 2018). Several studies carried out on agro-morphological characterization have successfully classified plantain cultivars in several countries (Amoutchi et al., 2020; Batte et al., 2018; Sivirihauma, et al., 2017b). Those studies indicated that a wide range of morphological traits could be a relevant tool for germplasm identification and selection.
Although many systematic studies continue to use morphological criteria, a great avenue in systematic studies of Musa has been launched using molecular data, particularly DNA analysis techniques such as RFLP, AFLP, RAPD, SSR, and diversity arrays technology (DArT) (Christelová et al., 2017; Ortiz and Swennen, 2014; Risterucci et al., 2009). However, these techniques are limited in differentiating plantain cultivars due to the low genotypical diversity within plantains (Crouch et al., 2000; Noyer et al., 2005), despite their high phenotypic diversity which is an important advantage for farmers who attempt to select varieties that suit their climatic conditions, markets and local habits (Dépigny et al., 2016).
Recent studies have been undertaken in Benin on the ethnobotany of Musa (Chabi et al., 2018), but the agro-morphological variability that characterizes plantain cultivars has not been investigated. The current study aims at inventorying all plantain cultivars available in small scale farming in southern Benin and classifying them using agro-morphological traits based on discriminating descriptors.
METHODOLOGY
Study area
Plantain accessions were collected between February and July 2019 in three districts (Athiémé, Tori-Bossito and Zê) on southern Benin (Figure 1). Those districts were identified within AVLANTO-Benin project as a great plantain production area of high diversity. This area is located in the sub-humid Guinean zone, between 6°25’and 7°30’N with a sub-equatorial climate featured by two dry and two rainy seasons. The average rainfall is 1200 mm while the mean annual temperature and the relative humidity vary between 25 and 29°C and between 69 and 97%, respectively (Hountondji et al., 2011).

Preliminary study and accessions collecting
In order to minimize duplication of accessions during plantain cultivars collecting, a census of all plantain producers in the three districts was made using the snowball principle, which is a chain sampling method consisting of targeting other participants based on the commendations of the previous ones. During this census, the local name of plantain cultivars produced by each producer, as well as the main phenotypic characteristics, were collected.
A focus group of 16 to 47 participants (Table 1) was organized per district with key informants represented by farmers who had been producing plantain for at least ten years (sampling method in Appendix 1). A formal local name and the phenotypic characteristics of the selected cultivars were agreed upon. For the validation of plantain cultivars local name by focus group, the principles of conducting a qualitative study of Cohen and Crabtree (2005) were strictly applied. Then, in the fields, twelve suckers of each accession were collected per district from plants bearing identifiable bunches. The suckers were labelled on the basis of data collected (local name, cultivar description and geographic coordinates of the site where suckers were collected from).

Installation of the collection
Accessions collected from the various districts were planted on an experimental plot of the Faculty of Agricultural Sciences of the University of Abomey-Calavi, located in Sékou (latitude 5° 18 'N and longitude 4° 09' W). The climate is sub-equatorial with two rainy seasons (a long season from March to June and a short season from September to November) and two dry seasons (from July to September and from November to March). The average annual rainfall varies between 800 and 1100 mm (Nangbe and Guidigbi, 2006).
The twelve suckers of each accession were planted on line with 2.5 × 2.5 m distances between rows and plants. Agricultural practices such as early and maintenance organic fertilization, irrigation, weed management, de-suckering and de-leafing were applied to ensure good plant development.
Characterization of accessions
Overall, sixteen plantain descriptors selected from the "MusaNet Taxonomic Advisory Group" list version September 2018 were used to characterize the accessions. These descriptors were selected from the work byAdheka et al. (2018)and included 5 quantitative (pseudostem height, number of fruits in the median hand of the bunch, fruit length, number of hands, and pedicel length) and 11 qualitative (bunch position, inflorescence type, pseudostem colour, bunch appearance, bract apex shape, fruit position, remains of flower relicts at fruit apex, fruit peel colour at maturity, pulp colour at maturity, bunch shape). Data were collected on 5 plants per accession at the appearance of ripening or yellowing signs on at least one finger of the first hand of the bunch.
The descriptors such as pseudostem height, bunch position, and inflorescence type were considered primary descriptors that allowed a rapid separation among plantain cultivars. Secondary descriptors were those that differentiate one cultivar from another within the main group. Pedicel length, fruit position and bunch shape are the rare descriptors that can only differentiate one cultivar from the others in the subgroup. This approach makes the description logical and faster; as it moves from general to specific traits, thus gradually reducing the number of descriptors to consider (Adheka et al., 2018).
Data analysis
A Factor Analysis of Mixed Data (FAMD) was performed to produce the data representation followed by a Hierarchical Ascending Classification (HAC) based on the "representative" factors from the FAMD. The number of morphological groups of plantain accessions was determined using the criterion of the partition with the highest inertia (Larmarange, 2016). Morphological characterization of different groups was performed based on quantitative discriminant descriptors from a Multivariate Analysis of Variance (MANOVA). The relationship between the quantitative discriminant variables was tested using the correlation circle. Statistical analyses were performed with R4.0.3 R Core Team (2020) software. The HCPC function of the factoextr package was used to determine the number of morpho-metric groups. Classification analysis was performed using the FAMD function from the Factoshiny package (Vaissie et al., 2021).
RESULTS
Inflorescence type (male rachis appearance)
Analysis of plantain diversity revealed the existence of three inflorescence types of the plantain subgroup in Benin (Figure 2). False horn plantains were predominant with 19 accessions, followed by French type plantains represented by 10 accessions and True horn plantains with 02 accessions (Table 2).


Variation of qualitative characteristics
Analysis of the variability of qualitative characters showed a high polymorphism of characters (Table 3). At full maturity, accessions showed mainly cylindrical (70.96%), loose (41.93%) or compact (25.80%) bunches with a slightly oblique position (83.87%) in relation to the pseudo-stem. A medium-green colour of the pseudostem was dominant with 67.74% among the accessions, followed by accessions with light-green pseudostem (29.04%). Only one accession "Aloga Kouélessi" stood out from the others with a purplish red pseudostem. A huge variability was observed in the shape of the bract apex with a predominance (61.28%) of the intermediate form. The obtuse and acute forms of the bract apex each had a proportion of 12.90%. Floral remnants were recorded at the tip of the fruit in the majority of accessions (83.87%) while only 16.13% of accessions had the base of the style prominent (without floral remnants). Most of the accessions in the collection had fruits with very little curvature (87.10%), with yellow pulp colour (74.20%) and light green peel at fruit maturity (93.56%).

Variation of quantitative characteristics
Analysis of variance of quantitative characteristics (Table 4) showed a highly significant difference between all variables (p<0.01). Thus, plantains grown in Benin were generally large (≥3 m) with an average height of 3.34 m. “Djangan ayawé” was the most giant accession (3.89 m) while hybrid L5449 was the shortest accession (2.89 m) in the collection. The number of hands in the bunches ranged from 2 to 11.67. The 'True horn' plantains identified in Benin ('Djangan ayawé' and 'Aloga alowé') have the specificity of bearing bunches with two hands. An average of 9.26 fruits were recorded on the median hand of the bunches of the accessions. On the median hand of the accessions 'Djangan djin yi', 'Djangan yi', 'Djangan agbavé', 'Gnivlan', 'Aloga kokolodjo' (P3-1), 'Aloga wéwé' (P2-2) and 'Aloga kokolodjo' (P2-1) the minimum number of fruits (6.67) was recorded. The maximum number of fruits (21.67) in the middle hand was observed in accession L5449. The average size of the inner fruit in the middle of the median hand was 22.90. The shortest fruits (16.83) were observed in accessions P2-13 and P3-7 named Pita 3 and the longest (35.67) in 'Djangan ayawé' and 'Aloga alowé'.

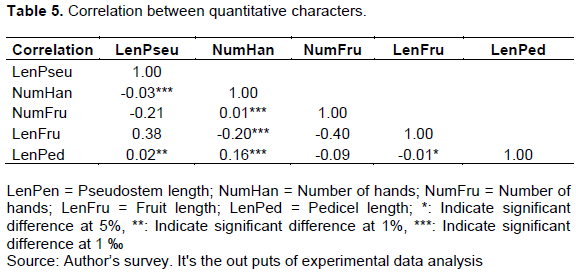
Correlation between quantitative characters
The Pearson correlation matrix (Table 5) showed significant correlations at p<0.05. The length of the plantain pseudostem (LenPseu) was significantly and positively correlated with the length of the pedicel (LenPed) (r = 0.02), as well as the number of hands (NumHan) with the number of fruits (NumFru) of the middle hand (r = 0.01) and the length of the pedicel (r = 0.16). Negative correlations were observed between length of plantain pseudostem and number of hands (r = -0.03), between fruit length (LenFru) and number of hands (r = -0.20) and between pedicel length and fruit length (r = -0.01).
Principal component analysis (PCA)
PCA showed that the first two axes contained up to 68% of the total variability. Pseudostem length (LenPseu), fruit length (LenFru) and pedicel length (LenPed) were positively correlated with PC1. Number of hands (NumHan) and pedicel length (LenPed) were positively correlated with PC2 (Figure 3).

Diversity and similarity among accessions
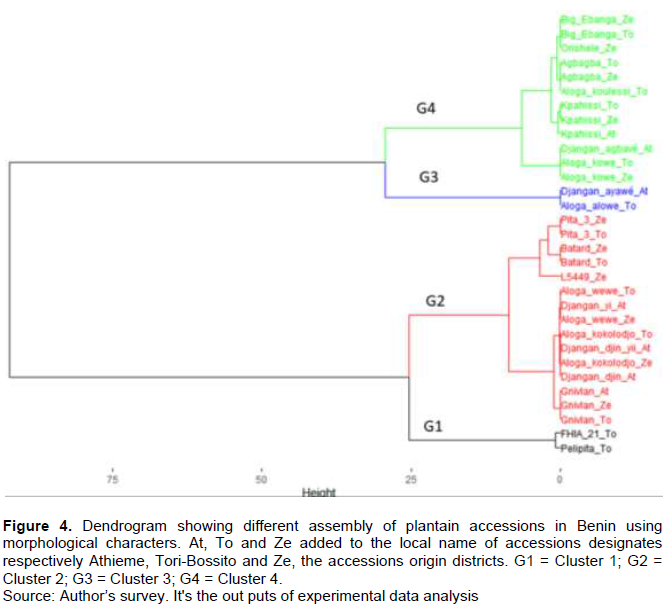
Hierarchical clustering of the mixed data showed that the first two axes express 93.63% variability in characters. The dendrogram analysis (Figure 4) showed the existing diversity and similarity among plantain accessions on the basis of agro-morphological characters. At a similarity index of 25%, four main clusters were identified and the number of accessions belonging to each cluster varied from 2 to 15. Thus, cluster 1 included two accessions (P2-11 and P2-12), as did cluster 3 (P2-4 and P1-5). Cluster 2 recorded the largest number of accessions including P2-6, P3-5, P1-7, P1-1, P3-1, P1-2, P2-1, P3-2, P1-3, P2-2, P3-6, P2-9, P3-9, P2-13, and P3-7 followed by cluster 4 with 12 accessions: P3-3, P2-3, P1-4, P1-6, P3-4, P2-7, P2-5, P3-8, P2-8, P3-11, P2-10, and P3-10. The analysis of multiple variance (MANOVA) revealed that the quantitative variables: length of the pseudo stem, number of hands, length of the fruit and length of the pedicel best explained the formation of the various clusters (Table 6). From the analysis of this table, it appears that the accessions of cluster 2 had the longest fruits (22.99±3.07). Individuals in clusters 3 were characterized by a long pseudo stem (3.85±0.06) with fruits having long pedicels (59.17±0.71). Accessions with the highest number of hands (7.53±2.52) were identified with cluster 4.

DISCUSSION
Main discriminating characters of plantains
The most discriminating descriptors that allowed quick separation between plantain cultivars include inflorescence type, pseudostem height, and bunch position (Adheka et al., 2018). Based on the typology of inflorescence the three types of inflorescence were recorded: False horn, True horn and French type plantains. False horn was the most represented with 19 accessions. This finding is contrary to the results from several works in which it came out that the French type was the most represented (Adheka et al., 2018; Hauser and Amougou, 2010). This may be due to high synonymy among False horn accessions (Hauser and Amougou, 2010). Subject to molecular characterization to elucidate this synonymy, it is clear that clusters 2 and 4 (Figure 4 and Table 2) included more accessions with names varying very little between sociolinguistic groups. These results are similar to that of the work by Sivirihauma et al. (2017b) in Ituri (DRC) where 14.4% French type, 44.5% False horn and 0.5% True horn were recorded in a collection.
Although banana pseudostem height is influenced by growing conditions and growth cycle (Adheka et al., 2018), the collection benefited an adequate monitoring that led to a good development of accessions. The average height of accessions was 3.34 m; which classifies accessions in Benin as giant. In fact, according to "MusaNet Taxonomic Advisory Group" Plantains of size ≤ 2 m are small; between 2.1 and 2.9 m are medium; while those ≥ 3 m are giant. This criterion is very discriminating and determinant, as it is involved in plant lodging, and consequently yield loss (Mustaffa and Kumar, 2012). In fact, plantains that are smaller than 3 m in height are more resistant to lodging (Chang et al., 2018). Only the accession (L5449) with 2.89 m fulfilled this condition, suggesting that all cultivars of plantain in Benin are highly exposed to lodging, especially in areas of cyclical high winds.
Three types of bunch orientation were recorded in the collection with a dominance of slightly oblique or sub-horizontal bunch accessions, but diversely represented in terms of inflorescence types. The True horn plantains 'Djangan ayawé' and 'Aloga alowé' and the French type Pelipita had oblique bunches to 45° or almost horizontal in relation to the pseudostem. Only the accessions FHIA 21 and L5449, all hybrids imported had vertically hanging bunches. Ortiz and Vuylsteke (1998), considered that the orientation of plantain bunches is an economically important trait. According to the authors, hanging bunches are more symmetrical than sub-horizontal, horizontal or erect bunches and are therefore more suitable for transport. Consequently, most plantain cultivars in Benin are not very prone to transport without the damage of fruits on the bunches.
Other agro-morphological characteristics of plantains
Analysis of diversity based on secondary and tertiary phenotypic characters revealed the formation of different clusters of plantain according to common similarities with accessions. In fact, some resemblances were recorded between the individuals of the same cluster. Despite those common criteria within a cluster, accessions held different local names depending on the sociolinguistic group. This result was also found by Ayenan et al. (2017)who indicated that vernacular names of plant genetic resources, generally vary from one ethnic group to another, from one village to another within the same ethnic area, and sometimes from one household to another within the same village. Similarly, accessions with the same name but belonging to different sociolinguistic groups were found in the same cluster. This is the case for accessions in cluster G2 named "Aloga kokolodjo" (P2-1 and P3-1) on one hand and "Gnivlan" (P2-6, P3-5 and P1-7) on the other. This may be due to redundancy of accessions in the plantain collection. According to Gonçalvès de Albuquerque et al. (2019), genetic redundancy presents a challenge for effective genetic resource management. Identification of genetic redundancy through molecular characterization becomes an effective approach to discriminate these accessions in the plantain collection (Koffi et al., 2019), in order to establish name equivalences between cultivars (Adheka et al., 2018), but also to reduce the cost of conservation (Horna et al., 2010).
This study also revealed that fruit curvature, fruit peel colour, and pulp colour were the lowest polymorphic characters. These characters were dominated by accessions bearing fruits with very little curvature (87.10%), yellow coloured pulp (74.20%) and light green peel (93.56%). This can be ascribed to the dominant traits selected by farmers for a long period of time, due to their acceptability by consumers who are key actors in the value chain in adopting genetic resources. These varietal preference criteria should be taken into account by any breeding program (Kinhoégbè et al., 2020) for plantain improvement in Benin. However, the work by Newilah et al. (2021) indicated farmers and consumers select mainly plantain cultivar based on the colour of the fruit pulp. In fact, cultivars bearing fruits with yellow pulp largely dominated plantain cultivars. Such preference contributes to the genetic erosion of white pulp cultivars. Surprisingly, some consumers associate the white pulp to the immaturity of plantain (Dadzie and Orchard, 1997). This result was confirmed by a recent study which revealed farmers’ behaviours considering orange/ yellowish pulp as an evidence of maturity of the fruit (Honfo et al., 2020).
Accessions "Djangan agbavé" (P1-4) and "Aloga kowé" (P2-3 and P3-3) showed the same agro-morphological characteristics with "Aloga kokolodjo" (P2-1 and P3-1), having carried simultaneously two bunches of fruit, which is not a stable character. This phenomenon can be assimilated to a genetic mutation and was also observed by Tenkouano (2000) and Odeigah (1997) who qualified it as a dichotomous inflorescence in plantain. Therefore, this trait cannot be considered since the characterization of genetic material must be based on stable, heritable traits that are consistently expressed and easily distinguished by the naked eye (Batte et al., 2018).
CONCLUSION
This paper investigated the diversity of plantain cultivars in southern Benin using agro-morphological descriptors. It came out that main descriptors such as type of inflorescence, height of the pseudosterm, position of the bunch made it easy to classifying those cultivars, based on some polymorphic traits. Also, the overall cultivars were giants, very prone to lodging; suggesting that new arenas of investigation should come into play to creating dwarf variety, and to promote the establishment of fence around plantain fields to break the wind velocity. The complementary of traits of cultivars has led to the formation of four distinctive clusters using criteria such as similarity in the length of the fruit, the pseudo-stem, the pedicel and the number of hands. This description highlighted the heterogeneity existing within plantain cultivars and consequently the agro-morphological diversity. Those findings are relevant in developing an effective strategy for varietal selection and conservation regarding plantain genetic resources. For this purpose, improving productivity and increasing plantain production in Benin should take into account accessions that combine interesting and economically profitable traits. A study on the genetic diversity of Benin plantains through molecular markers becomes a necessity, allowing to deepen the issue of taxonomy and synonymy for research improvement. Finally, a knowledge of the complete diversity of plantains in Africa could be possible by reconciling the results of the characterization of all the countries of Central and West Africa in order to progressively reach an agreement on the synonymy and uniqueness of accessions.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The research was fubded by the Dutch Organization for Scientific Research "NWO-WOTRO" within the framework of the AVLANTO-Benin project under Grant Agreement No: W 08.270.345. The "Agence Universitaire de la Francophonie (AUF)" also supported financially this study under Grant Agreement N°: DRAO-2017-U- S0531PRS20803.
REFERENCES
|
Ademiluyi BO (2013). Influence of number of sucker per plant on the growth , yield and yield components of Plantain ( Musa sp ) in Ado-Ekiti, Nigeria. Agricultural Science Research Journal 3(2):45-49. |
|
|
Adheka JG, Dhed'a DB, Karamura D, Blomme G, Swennen R, De Langhe E (2018). The morphological diversity of plantain in the Democratic Republic of Congo. Scientia Horticulturae 234:126-133. |
|
|
Amoutchi AI, N'nan-Alla O, Thiemele DEF (2020). Agro-morphological diversity of plantain accessions from different part of the world. International Journal of Biological and Chemical Sciences 14(4):1308-1321. |
|
|
Ayenan MAT, Danquah A, Ahoton LE, Ofori K (2017). Utilization and farmers' knowledge on pigeonpea diversity in Benin, West Africa. Journal of Ethnobiology and Ethnomedicine 13(37):1-18 |
|
|
Batte M, Mukiibi A, Swennen R, Uwimana B, Pocasangre L, Hovmalm HP, Geleta M, Ortiz R (2018). Suitability of existing Musa morphological descriptors to characterize East African highland 'matooke' bananas. Genetic Resources and Crop Evolution 65(2):645-657. |
|
|
Blomme G, Ploetz R, Jones D, De Langhe E, Price N, Gold C, Geering A, Viljoen A, Karamura D, Pillay M, Tinzaara W, Teycheney PY, Lepoint P, Karamura E, Buddenhagen I (2013). A historical overview of the appearance and spread of Musa pests and pathogens on the African continent: Highlighting the importance of clean Musa planting materials and quarantine measures. Annals of Applied Biology 162(1):4-26. |
|
|
Chabi MC, Dassou AG, Dossou-Aminon I, Ogouchoro D, Aman BO, Dansi A (2018). Banana and plantain production systems in Benin: Ethnobotanical investigation, varietal diversity, pests, and implications for better production. Journal of Ethnobiology and Ethnomedicine 14(78):1-20. |
|
|
Chang S, Wu Z, Sun W, Qiao L, Zeng Q, Zhang J, Shu H (2018). Factors Related to Physical Strength of Banana Pseudostem. Advances in Bioscience and Biotechnology 9(4):136-146. |
|
|
Christelová P, De Langhe E, H?ibová E, ?ížková J, Sardos J, Hušáková M, Van den houwe I, Sutanto A, Kepler AK, Swennen R, Roux N, Doležel J (2017). Molecular and cytological characterization of the global Musa germplasm collection provides insights into the treasure of banana diversity. Biodiversity and Conservation 26(4):801-824. |
|
|
Cohen DJ, Crabtree BF (2005). Evaluative Criteria for Qualitative Research in Health Care: Controversies and Recommendations. Annals of family medecine 6(4):331-9. |
|
|
Crouch HK, Crouch JH, Madsen S, Vuylsteke DR, Ortiz R (2000). Comparative analysis of phenotypic and genotypic diversity among plantain landraces (Musa spp., AAB). Theoretical and Applied Genetics 101(7):1056-1065. |
|
|
Dadzie BK, Orchard JE (1997). Routine post-harvest screening of banana/plantain hybrids: criteria and methods. INIBAP Technical Guidelines 2. In International Plant Genetic Resources Institute, Rome, Italy; International Network for the Improvement of Banana and Plantain, Montpellier, France; ACP-EU Technical Centre for Agricultural and Rural Cooperation, Wageningen, The Netherlands. |
|
|
Dépigny S, Lescot T, Achard R, Diouf O, Cote FX, Fonbah C, Sadom L, Tixier P (2016). Model-based benchmarking of the production potential of plantains (Musa spp., AAB): Application to five real plantain and four plantain-like hybrid varieties in Cameroon. Journal of Agricultural Science 155(6):888-901. |
|
|
Dochez C, Dusabe J, Tenkouano A, Ortiz R, Whyte J, de Waele D (2013). Screening Musa germplasm for resistance to burrowing nematode populations from Uganda. Genetic Resources and Crop Evolution 60(1):367-375. |
|
|
Folefack DP, Fongang Fouepe GH, Adamou KM, Ebongue JP, Bikoi A, Noupadja P (2017). Analysis of the plantain supply system of markets in the city of Douala. Journal of Economics and Sustainable Development 8(6):141-157. |
|
|
Gonçalvès de Albuquerque HY, de Oliveira JE, Brito AC, de Andrade LRB, do Carno CD, Morgante CV, Vieira EA, Moura EF, Faleiro FG (2019). Identification of duplicates in cassava germplasm banks based on single-nucleotide polymorphisms (SNPs). Scientia Agricola 76(4):328-336. |
|
|
Hauser S, Amougou D (2010). Plantain (Musa spp.) cropping systems of Southern Cameroon. Acta Horticulturae 879(2010):495-508. |
|
|
Honfo FG, Togbe EC, Ahohouendo AF, Ahohuendo BC (2020). Physical characteristics of some banana plantain cultivars (Musa spp.) consumed in Benin. Annales Université de Parakou, Série Sciences Naturelles et Agronomiques 10(2):17-22. |
|
|
Hountondji YC, De Longueville F, Ozer P (2011). Trends in extreme rainfall events in Benin (West Africa). 1960-2000., in The 1st International Conference on Energy. Environment And Climate Changes; Ho Chi Minh City. Vietnam 12 p. |
|
|
Horna D, Debouck D, Dumet D, Hanson J, Payne T, Sackville-Hamilton R, Sanchez I, Upadhyaya H, Van Den Houwe I (2010). Evaluating Cost-Effectiveness of Collection Management: Ex-situ Conservation of Plant Genetic Resources in the CG System. CGIAR, Montpellier, France. |
|
|
Kimunye J, Were E, Swennen R, Viljoen A, Mahuku G (2021). Sources of resistance to Pseudocercospora fijiensis, the cause of black Sigatoka in banana. Plant Pathology 70(7):1651-1664. |
|
|
Kinhoégbè G, Djèdatin G, Loko LEY, Agbo RI, Saxena RK, Varshney R, Agbangla C, Dansi A (2020). Agro-morphological characterization of pigeonpea (Cajanus cajan L) landraces grown in Benin: Implications for breeding and conservation. Journal of Plant Breeding and Crop Science 12(1):34-49. |
|
|
Koffi KGC, Pokou ND, Sie RS, Thiémélé DEF (2019). Genetic diversity and structure of plantain ( Musa sapientum L) landraces from Côte d'Ivoire using SSR markers. European Journal of Biotechnology and Bioscience 7(1):36-43. |
|
|
Larmarange J (2016). JLutils: Collection of R functions. R package version 1.11.0. |
|
|
Lescot T (2010). The genetic diversity of banana in figures. FruiTrop 177:48-52 |
|
|
Lescot T (2020). Banana genetic diversity. FruitTrop 269:92-96. |
|
|
Lokossou B, Affokpon A, Adjanohoun A, Dan CBS, Mensah GA (2012). Evaluation des variables de croissance et de développement du bananier plantain en systèmes de culture associée au Sud-Bénin. Bulletin de la Recherche Agronomique du Bénin (BRAB) pp. 10-17. |
|
|
Mohamed A, García-Martínez S, Loumerem M, Carbonell P, Ruiz JJ, Boubaker M (2019). Assessment of genetic diversity among local pea (Pisum sativum L.) accessions cultivated in the arid regions of Southern Tunisia using agro-morphological and SSR molecular markers. Genetic Resources and Crop Evolution 66(6):1189-1203. |
|
|
Mustaffa MM, Kumar V (2012). Banana production and productivity enhancement through spatial, water and nutrient management. Journal of Horticultural Sciences 7(1):1-28. |
|
|
Nangbe F, Guidigbi E (2006). Monographie de la commune de Allada. |
|
|
Newilah NG, Vepowo KC, Ngouno TA, Bouniol A, Rolland-Sabaté A, Meli Meli V, Lemoumoum JYS, Forsythe L, Dufour D, Fliedel G (2021). Analysis of consumer-oriented quality characteristics of raw and boiled plantains in Cameroon: implication for breeding. International Journal of Food Science and Technology 56(3):1135-1147. |
|
|
Noyer JL, Causse S, Tomekpe K, Bouet A, Baurens FC (2005). A new image of plantain diversity assessed by SSR, AFLP and MSAP markers. Genetica 124(1):61-69. |
|
|
Odah O, Aziadekey M, Tozo K, Akpavi S, Koukouma R, Guelly A, Kokou K, Assignon K, Akogo Y, Aidam A, Akpagana K, Kenny L, Ait-Oubahou A, Zinsou C, Gbeassor M (2013). La diversité génétique des bananiers plantains cultivés dans la zone Ouest de la Région des Plateaux au Togo. International Journal of Biological and Chemical Sciences 7(5):1910-1918. |
|
|
Odeigah PGC (1997). "Double bunching" in a plantain germplasm in Nigeria: A new genotype. Genetic Resources and Crop Evolution 44(1):1-3. |
|
|
Ortiz R, Swennen R (2014). From crossbreeding to biotechnology-facilitated improvement of banana and plantain. Biotechnology Advances 32(1):158-169. |
|
|
Ortiz R, Vuylsteke D (1998). Segregation of bunch orientation in plantain and banana hybrids. Euphytica 101(1):79-82. |
|
|
Risterucci AM, Hippolyte I, Perrier X, Xia L, Caig V, Evers M, Huttner E, Kilian A, Glaszmann JC (2009). Development and assessment of diversity arrays technology for high-throughput DNA analyses in musa. Theoretical and Applied Genetics 119(6):1093-1103. |
|
|
Saraswathi MS, Uma S, Vadivel E, Durai P, Siva SA, Rajagopal G, Sathiamoorthy S (2011). Diversity analysis in Indian cooking bananas (Musa spp) through morphotaxonomic and molecular characterisation. Acta Horticulturae 897:123-132. |
|
|
Schwartz SH (1992). Universals in the content and structure of values: Theoretical advances and empirical tests in 20 countries. In M. P. Zanna (Ed.), Advances in experimental social psychology, Academic P., M. P. Zanna, Ed. San Diego pp. 1-65. |
|
|
Sivirihauma C, Ocimati W, Vutseme L, Ntamwira J, Bahati L, Blomme G (2017a). Symptomless banana suckers sourced from Xanthomonas wilt infected fields are a viable alternative for seed within infected banana-based landscapes lacking access to clean planting materials. African Journal of Agricultural Research 12(31):2490-2498. |
|
|
Sivirihauma C, Ocimati W, Valimuzigha K, Karamura D, Adheka J, Ibanda B, Dhed'a B, Kamira M, Blomme G (2017b). Diversity and morphological characterization of Musa spp. in North Kivu and Ituri provinces, Eastern Democratic Republic of Congo. International Journal of Biodiversity and Conservation 9(10):292-305. |
|
|
Tenkouano A (2000). Persistence and horticultural value of inflorescence dichotomy in plantain. HortScience 35(5):933-936. |
|
|
Tenkouano A, Niéyidouba L, Agogbua J, Amah D, Swennen R, Traoré S, Thiemele D, Aby N, Kobenan K, Gnonhouri G, Yao N, Astin G, Sawadogo-Kabore S, Tarpaga V, Issa W, Lokossou B, Adjanohoun A, Amadji GL, Adangnitode S, Ortiz R (2019). Promising High-Yielding Tetraploid Plantain-Bred Hybrids in West Africa. International Journal of Agronomy 2019:1-8. |
|
|
Vaissie P, Monge A, Russon F (2021). Factoshiny: Perform Factorial Analysis from "FactoMineR" with a Shiny Application. R package version 2.4. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


