Agronomic potential of traditional sunflower spring varieties is low because their flowering and grain filling are often exposed to mid and end-season drought. To overcome this, new breeding strategy consisted of selecting varieties tolerant to winter cold in order to shift to autumn or early winter planting. Nowadays, ‘Ichraq’ is the only registered autumn variety in Morocco. The objective of this research was to evaluate various genotypes selected in different environments under autumn planting conditions. This germplasm was planted early during winter in 2014 at ‘Annoceur’, a mountain site known for its pronounced winter cold. Morphological, physiological, agronomic and technological parameters were considered for the germplasm assessment. Analysis of variance showed significant differences between genotypes for most of these parameters. Plantlet initial vigor average was 3.5 varying from 1 for genotype M32 to 5 for AN8. Leaf area average was 162 cm² varying from 25 to 375 cm² for genotypes M17 and AN34, respectively. Total chlorophyll content average was 43 mg/g, varying from 28 to 79 mg/g for genotypes K7 and M29, respectively. Number of days from sowing to flowering varied from 162 for genotype AN21 to 180 for genotypes M27 and M29. Mean seed yield per plant was 49 g, with a large variation from 8 to 110 g for M18 and K8, respectively. Mean seed oil content was 36%, ranging from 22% for M8 to 47% for K4. Plants exhibiting more performance than ‘Ichraq’ were selected to develop new sunflower germplasm suitable for autumn or early winter sowing.
Key words: Sunflower, genetic variability, autumn sowing, cold tolerance, physiological and agronomical traits.
Agricultural sector continues to dominate Morocco's economic activity. The rural population accounts for 40% of the total population. Thus, agriculture is an effective engine of economic growth and guaranteed food security. To upgrade and boost domestic agriculture, different strategies have been implemented during the Moroccan contemporary history. The latter being the Green Morocco Plan implemented since 2008. Owing to its importance in the cropping system and the food security challenge in vegetable oils, the oilseed is considered among the priority sectors. Since 2001, the year of oilseed sector reform, and until 2013, the year of signature of a sector program contract, sunflower has been the major annual oilseed crop grown in Morocco, with an average area of about 50000 ha and an average seed yield below 1 t/ha. Indeed, national seed oil production covers barely 2 to 3% of the overall needs of the edible oil in the country estimated at over 410.000 t. The gap is covered by importation which has negative repercussion on the economy and food security of the country (Nabloussi et al., 2015).
In Mediterranean area, sunflower traditionally sown in spring has limited productivity as it does not benefit from fall and/or winter precipitation, and it is often exposed to drought and high temperatures of mid and late cropping cycle. Such constraints coincide with periods of flowering and seed filling that are critical for determining seed productivity and seed oil content (Ouattar et al., 1992). Its cultivation is often secondary and is considered as catch crop, following early droughts or floods that affect autumn crops growing, mainly cereals. However, several studies have shown the benefits of early planting (autumn or early winter) in improving the seed yield and oil content in Morocco (Boujghagh, 1993; Gosset and Vear, 1995; Aboudrare et al., 2000), Spain (Gimeno et al., 1989) and France (Allinne et al., 2009). Early sowing, two to three months earlier than conventional sowing, induced a significant drop in temperature at planting and during early stages of vegetative growth (Allinne et al., 2009). Low temperature has an effect on maximum biological processes. It initiates a number of physiological alterations which make the plant to be more cold tolerant (Browse et al., 2001). Among the frequent metabolic changes, photosynthesis is the leading physiological process studied under low temperature conditions in various species including sunflower (Allinne et al., 2009), rice (Zhi-Hong et al., 2005), maize (Fryer et al., 1995) and Arabidopsis (Uemura et al., 1995). Photosynthetic variations are particularly characterized by changes in photochemical efficiency (Jompuk et al., 2005), in photoinhibition (Verheul et al., 1995) and reaction to photooxidation (Wise et al., 1995). The characterization and evaluation of sunflower genotypes adapted to low temperature conditions, during early vegetative growth stages, requires analyzing the impact of such conditions on the physiological processes associated with initial seedling vigor and plant cold tolerance. Agronomic, morphological, physiological, technological and biochemical attributes could be taken as valuable criteria to identify and select genotypes adapted to winter cold conditions. Early sowing of sunflower in Moroccan environment was envisaged such as mid-season and terminal drought avoidance strategy. However, this implies early stages of crop development are exposed to winter cold which could be harmful to sunflower as it is known as a spring crop sensitive to low temperatures, especially during early growth stages. Therefore, breeding new varieties tolerant to low temperatures, and thus adapted to autumn or early winter planting, is necessary to adopt this strategy (Allinne et al., 2009).
Nowadays, “Ichraq” is the only autumn variety registered in the Moroccan Official Catalogue (Nabloussi et al., 2008). It is a late maturing, drought and winter cold tolerant and combines good seed yield and high seed oil content. Current research continues to develop new sunflower populations, resistant (or tolerant) to winter cold and agronomically performant, which would be the basis for selection of new improved varieties better than the variety “Ichraq”. Thus, the present work aimed to evaluate new sunflower genetic materials for agro-morphological, physiological and technological traits under early winter planting conditions.
Plant material
The plant material used in this study consisted of 46 sunflower genotypes including ‘Ichraq’, the first and only one autumn variety, considered as check, and 45 individual selected plants derived from ‘Ichraq’. As this latter is a population variety (Nabloussi et al., 2008), there was opportunity to select individual plants (PS) in order to release new germplasm that will be more performant than ‘Ichraq’. The 45 PS were selected under various environmental conditions, in early winter planting, for their vigor, habit and agro-morphological performances. Most of the 45 genotypes selected, being more performant than ‘Ichraq’, were phenotypically different from each other. The main selection criteria were earliness, reduced number of leaves per plant, leaves hairiness, high seed oil content and absence of diseases.
Methods
The 46 genotypes were planted on 2 January 2014 at the INRA experimental station located at ‘Annoceur’, mountainous area known for its rough winter cold. It is located 50 km from Fez city, in the north of Morocco, at an altitude of 1350 m (33°41'05.2"N 4°51'19.9"W).
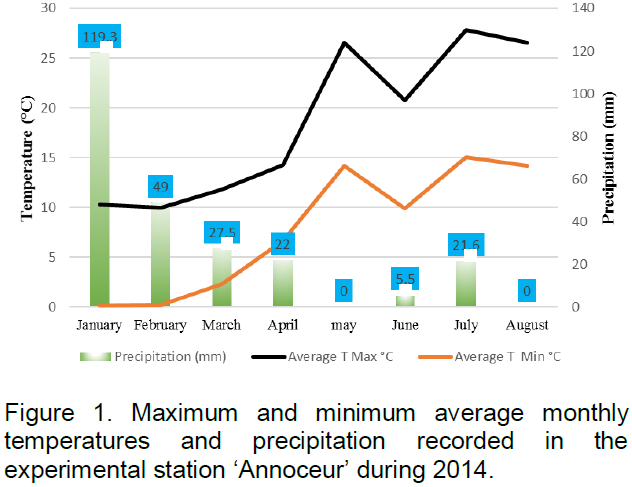
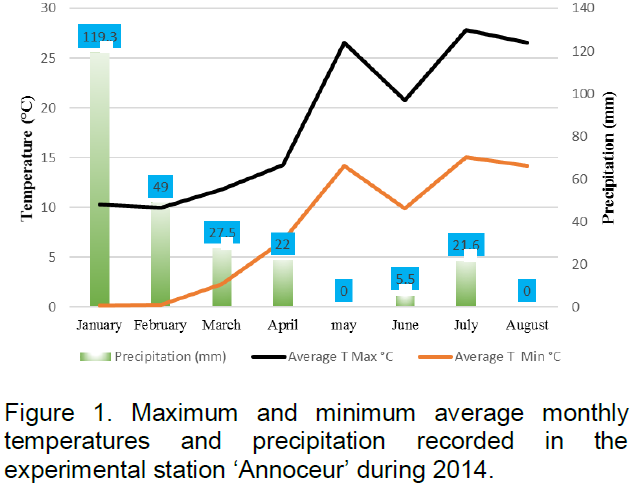
During the cropping cycle, the minimum temperature was -5°C, registered on January and February whilst the maximum temperature was 37°C, recorded on May. On the other hand, cumulative rainfall was around 245 mm, and the rainiest month was January with about 119 mm, ensuring good emergence and early seedling establishment. Figure 1 shows maximum and minimum average monthly temperatures and rainfall, characterizing the experiment environment. Daily data were gathered from the meteorological station located in ‘Annoceur’ experimental station.
The experiment was conducted under rainfed conditions following a randomized complete blocks with two replications. Each genotype was grown in a single 5 m-length row, and inter and intra row spacing were 60 and 30 cm, respectively. Initial N-P-K fertilization was 80-80-30 units, respectively, followed by cover N fertilization with two inputs of 40 units, one at stem elongation stage and the other at flowering stage. No phytosanitary treatment was applied.
Morphological, phenological, physiological, agronomic and technological parameters were studied. During plant vegetative growth, plant height (cm) was measured from the ground to the top of the plant at the maturity, and growth rate (cm/d) was determined at stem level, using a graduated ruler. Collar diameter (mm) was measured using a caliper. Initial vigor of young seedlings was evaluated at two leaves stage according to a grading scale from 1 (weakest vigour) to 5 (strongest vigour). Number of leaves per plant and number of branches per plant were determined at full flowering. Leaf area (LA, cm2) was also measured at full flowering using the following formula (Kiani, 2007): LA = Length x Width x 0.7. Flowering time of each genotype was determined by counting the number of days between planting date and the date when 50% of plants of this genotype have flowered. Chlorophyll content (mg/g) was calculated according to the method of Billore and Mall (1975). The optical density (OD) of all the supernatant obtained was measured in a spectrophotometer at 645 and 663 nm. The concentrations of chlorophyll pigments are given by the following formulas:
CHL A = 12.7 (OD 663) – 2.69 (DO 645)
CHL B = 22.9 (OD 645) – 4.56 (DO 663)
At maturity, head diameter and head aborted diameter, expressed in cm, were measured using graduate ruler. After harvest, seed yield per plant (g) was determined by weighting total plant seeds, using a precision balance. Also, its components (number of seeds per propeller and 1000 seeds weight) were determined. Seed oil content was determined using NMR method (Oxford 4000). This method of quantitative determination is reliable, fast and easy to implement. It analyzes directly on whole seed, without preparation or destruction of biological material.
From each genotype and each block, four plants were taken randomly to be used for all the measurements considered in this study both in field and laboratory. Descriptive analysis of gathered data, analysis of variance and analysis of correlation were performed using different procedures of SAS program. Duncan’s new multiple range test was applied to compare genotypes means. This enabled us to distinguish which genotypes were significantly different from others and, thus, to select those interesting genotypes according to an established threshold. Selection threshold was fixed on the basis of nature of parameters studied and their relation with abiotic stresses (cold and drought) and agronomic performance, overall average of each parameter and check performance. Furthermore, Agglomerative Hierarchical Clustering (AHC), created by XLSTAT statistical software, was performed to group homogeneous genotypes.
Morphological parameters
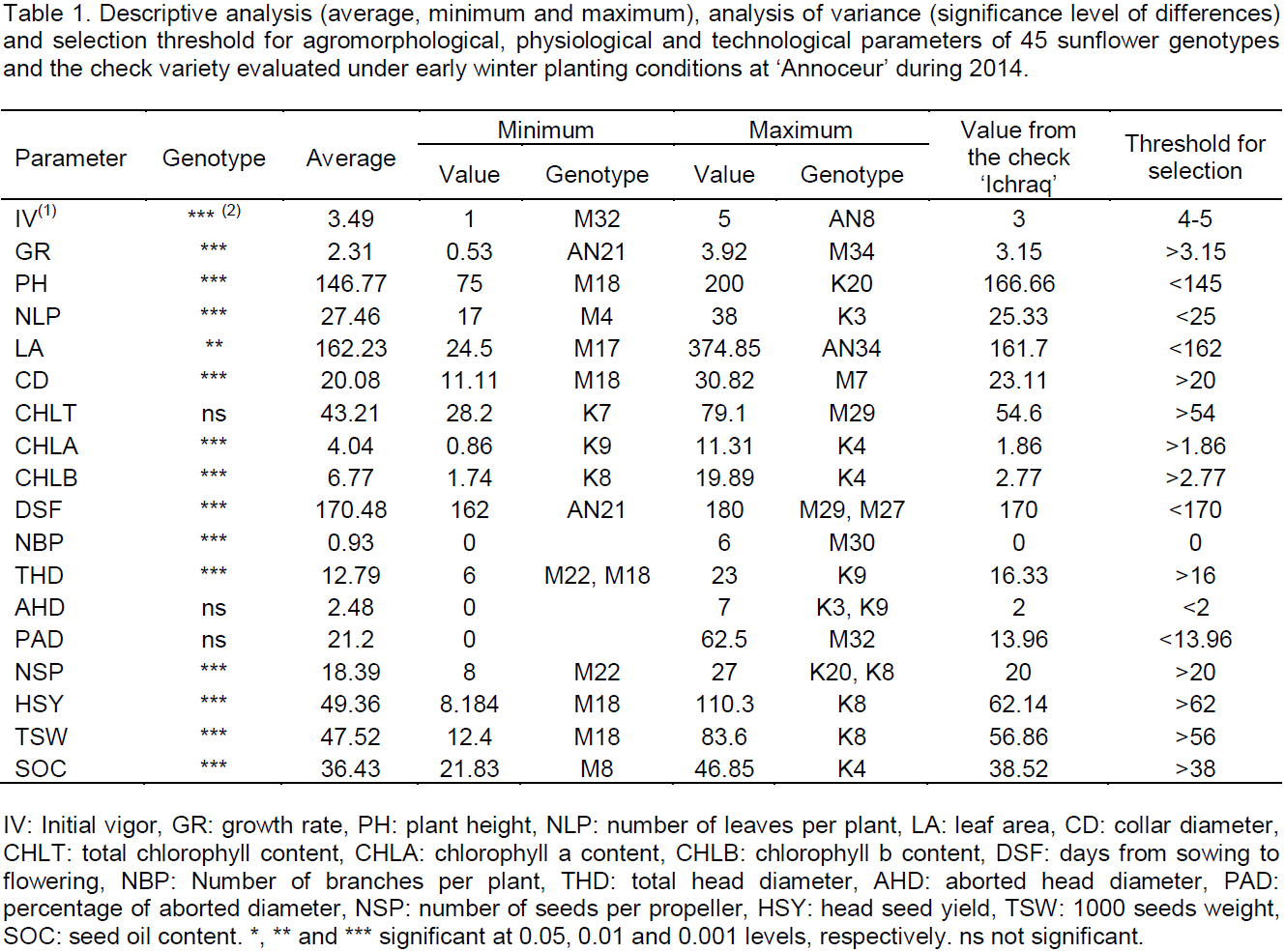
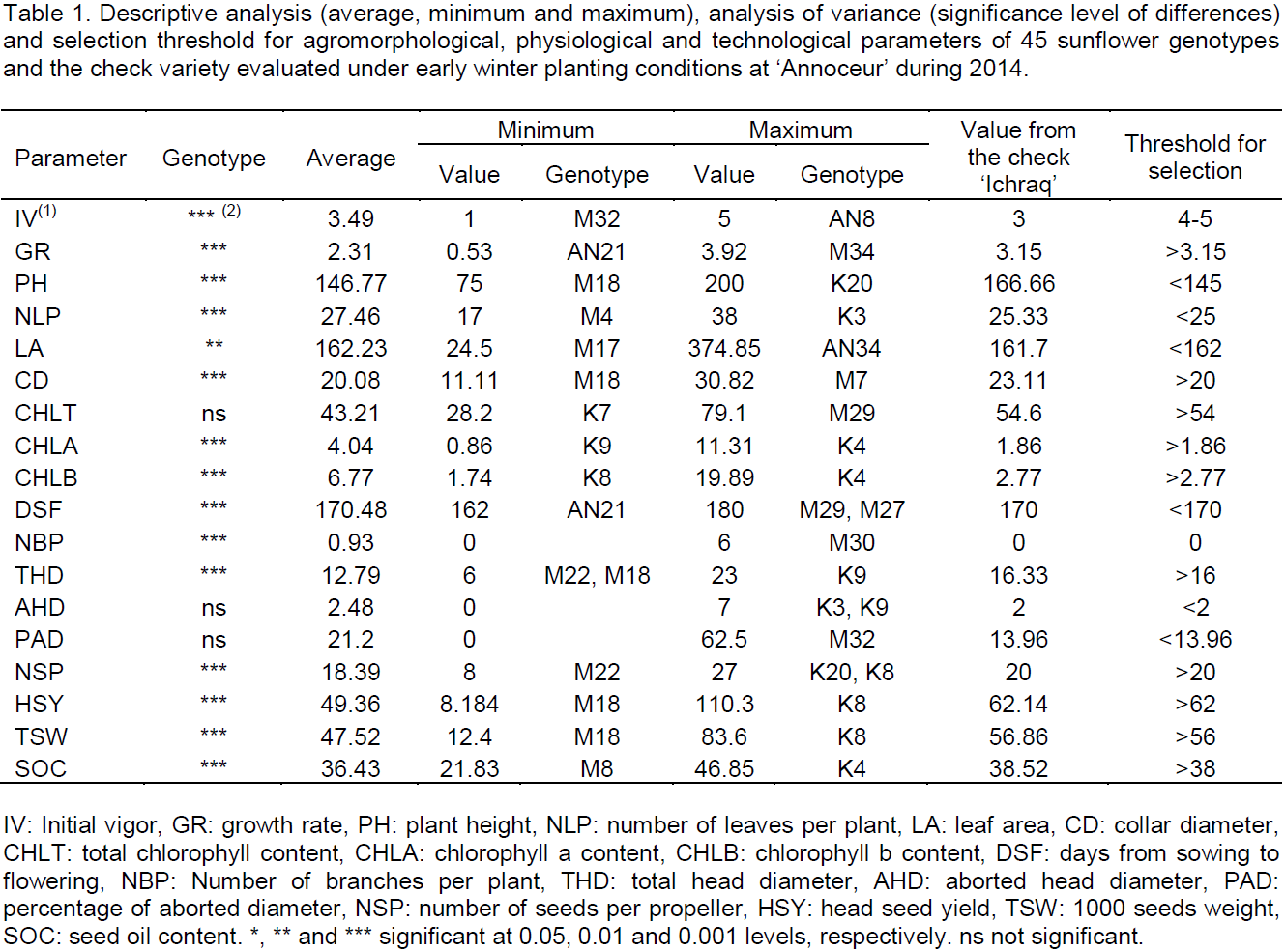
Analysis of variance showed there were significant differences (P < 0.001) between the 46 genotypes for all studied parameters (Table 1). Initial vigor of young seedlings varied from 1 for genotype M32 to 5 for genotype AN8, with an average of about 3.5, higher than ‘Ichraq’ vigor (3). In many studies, seedling and plantlet initial vigor was found as a good selection criterion correlated with adaptation and performance of genotypes under environmental abiotic stresses (Foolad and Lin, 2001). In the present work, all genotypes having an initial vigor of 4 or 5 were selected for further evaluation and germplasm improvement. For growth rate, the overall mean was 2.31 cm/d, with a minimum of 0.53 cm/d, registered for genotype AN21 and a maximum of 3.92 cm/d for genotype M34, slightly higher than that of the check, which was 3.15 cm/d (Table 1). Genotypes having growth rate higher than that of the check will be selected. The average plant height was 147 cm, with a variation from 75 to 200 cm for M18 and K20, respectively. Plant height of the check was about 167 cm. It was reported that very high plants were often susceptible to lodging and late drought (Sposaro et al., 2008). Thus, plants with a height less than the observed average (< 145 cm) could be interesting for selection. Number of leaves per plant varied from 17 for M4 to 38 for K3, with an average of 27.5 leaves per plant. The check had 25 leaves per plant. The average leaf area was 162 cm², which is equal to the check value. The genotypes M17 and AN34 exhibited the extreme values: 24.5 and 374.85 cm², respectively. Specific leaf area was found to be genetically associated with cold tolerance in sunflower (Allinne et al., 2009). Furthermore, elevated number of leaves per plant and high leaf area are correlated with high plant transpiration (Romero-Aranda et al., 2001). Thus, this study aimed to select those plants having less than 25 leaves and a leaf area less than 162 cm². Regarding collar diameter, genotype M7 exhibited the strongest value which was about 31 mm, whilst genotype M18 showed the lowest value which was 11 mm. The overall mean value was 20 mm and the check value was 23 mm. Like initial vigor, collar diameter is an indicator of good adaptation under stressed environments (Liua et al., 2012a). Therefore, all the genotypes exhibiting a collar diameter more than the observed average (20 mm) could be selected for further evaluation. Among the 46 studied genotypes, 27, including ‘Ichraq’, the check variety, had no branching, while 19 were branched, with a number of branches per plant varying from one to six. Genotype M30 was the most branched, having six branches per plant. The overall average was 0.93. Sunflower branching is an indicator of plants susceptibility to cold conditions (Alba et al., 2010). The plants selected for further evaluation and new germplasm constitution should have no branching.
Physiological parameters
Analysis of variance revealed significant effect of genotype on flowering earliness, chlorophyll a content and chlorophyll b content (P < 0.001), and non-significant effect on total chlorophyll content (Table 1). However, a large variation was observed, ranging from 28 mg/g for genotype K7 to 79 mg/g for genotype M29. The average total chlorophyll content was 43.21 mg/g, while the content concerning the check variety was 54.6 mg/g (Table 1). Genotypes maintaining high chlorophyll content under abiotic stresses, like drought or cold, exhibit tolerance to such stresses (Yang et al., 2015). In sunflower, chlorophyll content is genetically associated with cold tolerance and, thus, could be suggested as selection criterion for cold tolerance in breeding programs (Allinne et al., 2009). All genotypes having total chlorophyll content higher than that of the check will be selected. Regarding chlorophyll a and b contents, the genotype K4 exhibited the highest values for both, 11.3 and 19.8 mg/g, respectively. The lowest contents were 0.86 mg/g, registered in genotype K9, and 1.74 mg/g, registered in genotype K8, for chlorophyll a and b, respectively. The check variety had 1.86 and 2.77 mg/g for these parameters, respectively. Vegetative period before flowering was too long, with an average duration exceeding 170 days from sowing date to flowering date. It ranged from 162 days for genotype AN21 to 180 days for genotypes M27 and M29. The check variety has bloomed in 170 days after sowing. Flowering earliness is a desired character in environments under terminal drought stress (Ribot et al., 2012). Thus, genotypes having a sowing-flowering period shorter than that of the check will be selected.
Agronomic and technological parameters
Analysis of variance showed there were significant differences (P<0.001) between the studied genotypes for all agronomic and technological parameters, excepted aborted head diameter (AHD) and percentage of aborted diameter (PAD) (Table 1). However, one could observe some variation among genotypes for these parameters (Table 1). Most of the evaluated genotypes had no AHD, and among the few ones having AHD, genotypes K3 and K9 exhibited the largest value (7 cm). The overall average AHD was about 2.5 cm. The overall average PAD was about 21%, ranging from 0%, for most of the genotypes, to more than 62%, for genotype M32. All genotypes exhibiting some AHD should be discarded from the selected population as aborted sunflower head is an indicator of plant susceptibility to cold (Hladni et al., 2010). Average total head diameter (THD) was 12.8 cm, ranging from 6 cm, for genotypes M22 and M18, to 23 cm, for genotype K9. The check variety ‘Ichraq’ had a THD of 16 cm, an AHD of 2 cm and a PAD of 14%. A large range was observed for number of seeds per propeller, from 8 in genotype M22 to 27 in genotypes K8 and K20. The check variety had a number of 20 seeds per propeller.
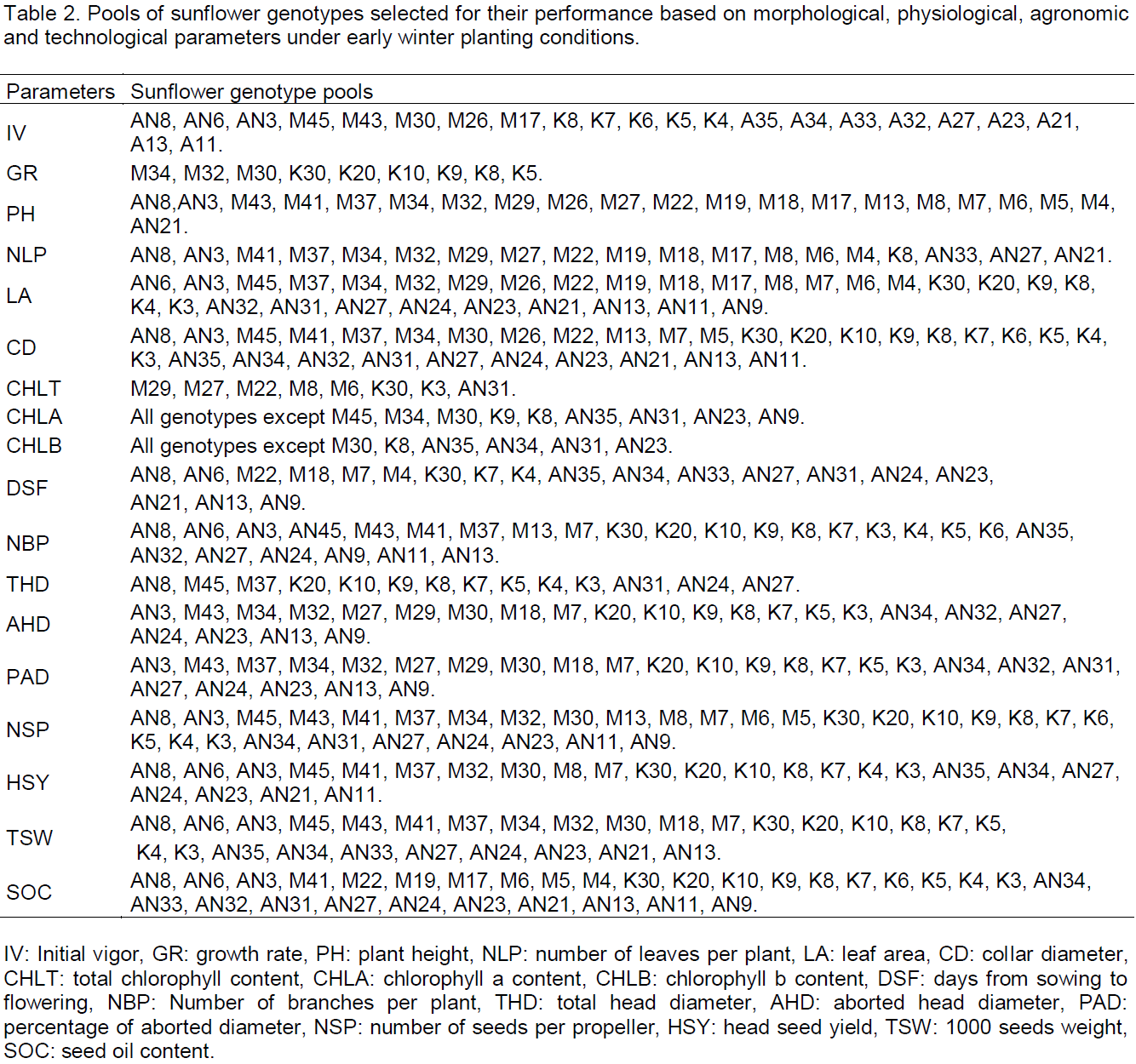
Regarding seed yield per head, the overall mean was slightly higher than 49 g, and a large range was found, from 8 g in genotype M18 to 110 g in genotype K8, which is much higher than head seed yield of the check (62 g). Thousand seed weight (TSW) ranged from 12.4 g in genotype M18 to 83.6 g in genotype K8, and the average was 47.52 g. TSW of the check was about 57 g. Total head diameter, number of seeds per propeller, single head seed yield and TSW are components of seed yield which are correlated with this latter, and thus could be considered as selection criteria for seed yield breeding (Yasin and Singh, 2010). In this study, all those genotypes showing values higher than those of the check were selected. Finally, seed oil content (SOC) fluctuated from 21.80% in genotype M8 to 46.85% in genotype K4, and had a mean value of 36.43%. The check ‘Ichraq’ had a SOC of 38.52%, which was slightly higher than the overall average. Genotypes with SOC exceeding that of the check were selected. Table 2 shows the pools of genotypes selected, according to described threshold for each of studied parameters.
Pearce (1999) subdivided the plants into three categories according to their tolerance to cold and ability to adapt to low temperatures. Plants are susceptible to low temperatures and suffer damage as early as 12°C, plants are tolerant to low positive temperatures and plants capable of acclimatizing to survive under temperatures below zero degree. Xin and Browse (2000) showed that there are many physiological mechanisms that allow plants to better withstand severe stress (temperatures below zero) after a long time at low temperature (acclimatization). Many studies have shown low temperature had direct effects on cells (Pearce, 1999), on seed germination (Durr et al., 2001), on photochemical reactions of photosynthesis and carbon fixation (Liua et al., 2012b). Likewise, cold causes reduction of cell water content (Kacperska, 2004).
These findings have shown there was a genetic diversity among the sunflower genotypes evaluated for most of the studied parameters. In all cases, these genotypes were compared with the check variety ‘Ichraq’. This study allowed us to identify and select genotypes more interesting than the check for morphological, physiological, agronomic and technological parameters under winter early planting conditions. Globally, taking into account all these parameters, the genotypes AN8, AN27, AN23, AN21, AN24, K30, K20, K10, K8, K7 and K4 seemed to be performant and promising. After confirming their performance in further seasons, they could be useful for intercrossing to develop a new variety more performant and more tolerant to winter cold than ‘Ichraq’, the only one autumn variety ever registered in Morocco.
On the other hand, molecular analysis to better understand the genetic control of agronomic, morphological and physiological traits associated with cold tolerance would be of a great interest for autumn sunflower breeding programs. Allinne et al. (2009) studied the genetic control of physiological traits associated to cold tolerance in sunflower under early sowing conditions and found that several putative genomic regions were involved in the variation of such physiological traits. Some SSR markers are associated with major QTLs for cold tolerance, such as ORS331_2 for cell membrane stability, ORS1040 for chlorophyll content and ORS1144 and ORS1146 for osmotic potential.
Genotypes hierarchical grouping
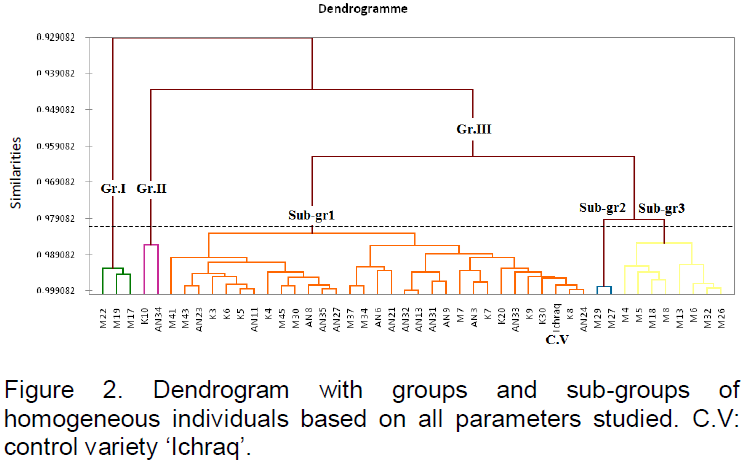
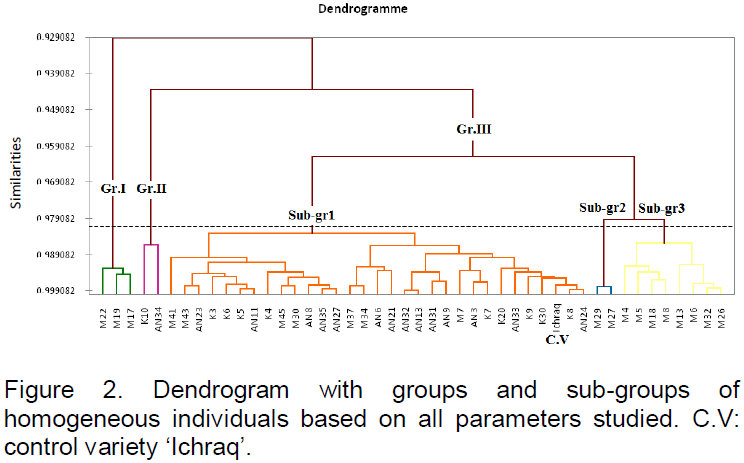
Agglomerative hierarchical clustering (AHC) allowed obtaining the dendrogram showed in Figure 2. It presents AHC results as similarity index (Pearson correlation) in groups of homogeneous genotypes on the basis of all studied parameters. Three distinct groups with a very high level of similarity could be identified. The first group contains three genotypes, M17, M19 and M22, which are similar for the parameters PH, NLP, LA and SOC. The second group is formed by only two genotypes, K10 and AN34, which are comparable for CD, AHD, PAD, NSP, HSY, TSW and SOC. The third group contains the rest of the studied genotypes and is divided into three homogeneous subgroups on the basis of all the parameters, with a similarity index of about 0.98. The first homogeneous subgroup constitutes 31 genotypes, including the control variety ‘Ichraq’. The second subgroup contains only two genotypes, M29 and M27, comparable for PH, NLP, CHLT, AHD and PAD. The last subgroup contains the genotypes M4, M5, M18, M13, M6, M32 and M26, with a similarity index of about 0.99, comparable for PH, NLP, LSA and NGP.
Previous studies has also shown sunflower genotypes hierarchical clustering and usefulness of genotypes grouped in the same pool, according to agronomic, phenological and technological character, for breeding programs in various countries in the world. Among these studies, Poletine et al. (2012) evaluated 16 sunflower genotypes in Brazil, throughout analysis of seven agronomic and morphological characters, and could classify them into five distinct groups. Another work carried out at Tunisia classified 73 sunflower lines and 7 hybrids (checks) into 5 groups and 8 sub-groups, based on morphological and phenological parameters, with a similarity coefficient of 90% (Khoufi et al., 2013).
Likewise, Ruždik et al. (2015), based on agronomic parameters, had classified 20 sunflower varieties into four groups in their study in Republic of Macedonia. Classifying thirteen sunflower genotypes in Libya by UPGMA procedure, on morphological basis, Mahmoud et al. (2012) had found two major groups. Similar results were described by Kholghi et al. (2011) in Iran and Punitha et al. (2010) in Coimbatore. All these homogeneous groups and subgroups carried out in these studies were useful for corresponding local sunflower breeding programs.
In this study, the homogeneous groups and subgroups could be taken as genetic pools to be used in autumn sunflower breeding program. Each group should be improved and multiplied apart before any crossing or intercrossing with the other groups.
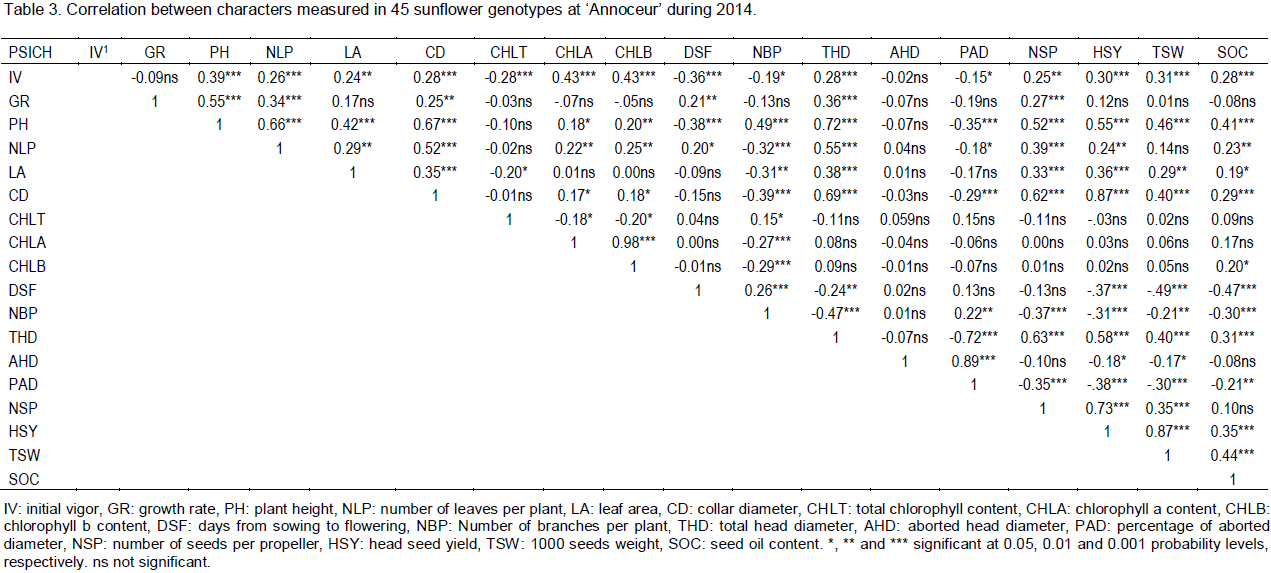
Relationship between characters
Table 3 shows correlation coefficients between the different parameters studied along with their significance level. There are significant correlations between plant seed yield (HSY) and other related parameters. Such correlations are either positive, with SOC, TSW, NSP, THD, CD, LA, PH and IV, or negative, with PAD, DSF, NBP and AHD. These results indicate that non-branched and early flowering plants with reduced aborted head diameter could be more productive than branched and late plants. However, the strongest correlations were observed between HSY with TSW (0.87, P<0.001) and NSP (0.73, P<0.001), indicating that TSW and NSP are the most important seed yield components, and thus could be taken as selection criteria to improve sunflower productivity under autumn early planting conditions. On the other hand, seed oil content (SOC) is significant and positively correlated with IV, PH, CD, THD, HSY and TSW, and significant and negatively correlated with DSF and NBP. Similar results on the positive correlation between SOC and TSW was also found by Joksimovic et al. (2004), Mijić et al. (2009), Kaya et al. (2009) and Anandhan et al. (2010). The highest correlations were observed between SOC and DSF (-0.47, P<0.001) and TSW (0.44, P<0.001), indicating that early flowering genotypes with large seeds could be characterized by high seed oil content. Interestingly, one could observe the significant and positive correlation between HSY and SOC (0.35, P<0.001). This will allow simultaneous breeding and selecting for both characters. Early flowering, large seeds and high seeds number per head could be valuable selection indices to bred high seed oil yielding germplasm under autumn sunflower planting conditions.
In conclusion, a large variability was observed between the evaluated genotypes for most of the traits studied. Genotypes having expressed more performance than the check ‘Ichraq’ in terms of seed yield, oil content and flowering earliness should be confirmed in further studies under autumn planting conditions in other additional environments. They could be considered as valuable genetic stock for autumn sunflower breeding program. The clustering homogeneous groups should be taken as genetic pools to be separately multiplied and thereafter intercrossed with each other to develop a new cold tolerant germplasm with better agronomic and technological performance than the check variety ‘Ichraq’. Specific molecular markers such as SSR should be used to confirm these homogeneous groups and to be considered thereafter in autumn sunflower breeding programs.
To bred autumn sunflower for both high seed yield and high seed oil content, the most relevant selection criteria would be early flowering, large seed size and high seeds number per head.