Full Length Research Paper

ABSTRACT
Drought is the most significant environmental calamity on sorghum in Ethiopia and hence improving yield under drought is a major goal of plant breeding. This study was designed to introgress drought tolerant genes into adapted varieties through marker-assisted backcrossing and select based on tolerance indices. Sixty-one backcrossed lines and along with their nine parental lines were evaluated under full-irrigation and water-limited condition in Alpha lattice design with three replications. Yield-based drought tolerance indices including stress tolerance index (STI), mean relative performance (MRP), geometric mean productivity (GMP), harmonic mean (HM), mean productivity(MP), tolerance index (TOL), stress susceptible index(SSI), yield stability index (YSI) and yield index (YI)were calculated based on yield obtained from the two moisture regimes. Results showed that genotypes differed significantly in yield and their indices. Mean grain yields that varied widely in stressed (1.1 to 4.42 t ha-1) and full-irrigation (2.25 to 5.71 t ha-1) were 1.93 and 3.7 t ha-1, respectively. Of the backcrossed lines, four (BC2F3_ETSC_16258,BC2F3_ETSC_16216, BC2F3_ETSC_16257, and BC2F3_ETSC_16213) were top yielding in stressed conditions with values of 4.42, 3.5, 3.1, and 2.83 t ha-1, respectively. These progenies also showed consistently higher values of STI, MRP, GMP, HM, MP, YSI, and YI and lower values of SSI and TOL indicating less sensitive to stress. The correlation and principal component analyses also revealed STI, MRP, GMP, HM, MP and YI showed highly significant positive correlation among themselves and yield in both environments, indicating their suitability for identifying superior genotypes. Overall, STI, MRP, GMP, HM and MP indices can be efficiently exploited to screen drought tolerance or superior genotype(s) under both moisture conditions.
Key words: Coefficient of correlation, drought tolerance indices, principal component, clusters analysis.
INTRODUCTION
Sorghum, Sorghum bicolor (L.) Moench is an important cereal crop in many parts of the world grown for food, feed, and industrial purposes (Reddy, 2017; Visarada and Aruna, 2019). It is one of the most important dry land food crops grown in marginal lands and dietary food for more than half a billion poor and most food insecure people living in the sub-tropical and semi-arid regions of Africa and Asia (FAO, 2017).Sorghum is produced in intensive and commercialized in developed world with average yields of 3-5 t ha-1 largely used for feed, while, in the developing countries, it is grown in low-input, extensive production systems, with productivity of being 1 t ha-1 mostly for food (Kumar, 2016; Reddy, 2017). Ethiopia is the sixth largest producer of sorghum in the world after USA, Nigeria, Mexico, Sudan and India and the third in Africa behind Nigeria and Sudan (FAO, 2017) with sorghum contributing 16.89% of the total annual cereal grains production occupying approximately 1.9 million ha of land (CSA, 2018). Sorghum takes the third largest share of all cereals grown in Ethiopia next to tef [Eragrostistef (Zucc.) Trotter] and maize (Zea mays L.) be it in hectare or volume of total annual national production (CSA, 2018). It provides more than one third of the cereal diet and acts as a principal source of food, feed, income and beverages for millions of the resource-poor people (MoA, 2018) dwelling in marginal areas where drought is the primary production constraint (Amelework et al., 2015; Mera, 2018; Teshome and Zhang, 2019; Wagaw, 2019).
Despite the potential and multitude uses of sorghum, however, the full genetic potential of the crop cannot be harnessed particularly in tropical and sub-tropical Africa including Ethiopia because of limitations simultaneously imposed by attacks from biotic and abiotic constraints. Of the abiotic constraints, drought is an important limiting factor for sorghum production in most parts of the world including Ethiopia, ultimately influencing yield and quality (Harris et al., 2007; Kassahun et al., 2010; Sabadin et al., 2012; Reddy et al., 2014; Madhusudhana, 2015; Amelework et al., 2015, Sory et al., 2017; Mera, 2018; Teshome and Zhang, 2019; Wagaw, 2019). Yield loss due to drought in the tropics alone exceeds 17% of well-watered production, reaching up to 60% in severely affected regions (Ribaut et al., 2002; Sharma and Lavanya, 2002). In Ethiopia, where more than 50% of the total area is semi-arid, insufficient, unevenly distributed, and unpredictable rainfall is usually experienced in drier parts of the country (Amelework et al., 2015; Mera, 2018; Teshome and Zhang, 2019).It is manifested by delay in onset, dry spell after sowing, drought during critical crop stage and too early stop. It is frequently observed that drought is occurring at more frequent intervals-every two years during recent years. For instance, between 1960 and 1990 there were six droughts in the country, but between 1990 and 2014 there were nine droughts (Mera, 2018) caused up to complete annihilation of sorghum and other crops affecting millions of people. This showed that climate change makes increasing production much more challenging. Recent reports also declare that the intensity and frequency of droughts are expected to increase, resulting in decreased food production and food security and increased vulnerability of the crop to drought (Bates et al., 2008; Wassmann et al., 2009; Mera, 2018; Teshome and Zhang, 2019).
Among the drought management strategies, genetic manipulation of the crop to improve tolerance is preferred because of its sustainability and feasibility particularly to the resource-poor (Singh, 2002; Keneni, 2007).Breeding for drought-tolerant crops largely depends on the availability of the genetic resources for tolerance, reliable screening techniques, identification of genetic components of tolerance (Blum, 2011), successful genetic manipulation of the desired genetic backgrounds, and ultimate development of drought-tolerant cultivars with acceptable agronomic and quality-related traits (Araus and Cairns, 2014). The relative yield performance of genotypes under drought stressed and non-stressed environments can be used as an indicator to identify drought resistant varieties in breeding program for drought prone areas (Raman et al., 2012; Mohammadi, 2016). Based on their comparative yield performance in stress and non-stress environments genotypes were categorized in four groups; genotypes with high performance under both moisture regimes (group A), high yield in non-stress conditions (group B), high yield in stress conditions (group C), and low yield under both moisture regimes (group D) (Fernandez, 1992). In this regard, several drought indices that are based on drought resistance or susceptibility of genotypes have been suggested and computed between yield under stress and optimal conditions. Drought indices which provide a measure of drought based on loss of yield under drought conditions in comparison to normal conditions have been used for screening drought tolerant genotypes.
Thus, many authors have been reported that the relative merits of different indices for screening of genotypes to drought based on their comparative yield performance in stress and non-stress environments. These include; stress tolerance index (STI) and geometric mean productivity (GMP) (Fernandez 1992), stress susceptibility index (SSI) (Fischer and Maurer, 1978), tolerance index (TOL) (Hossain et al., 1990), mean productivity (MP) (Rosielle and Hamblin, 1981), yield index (YI) (Gavuzzi et al., 1997), yield stability index(YSI) (Bouslama and Schapaugh, 1984), harmonic mean (HM) (Schneider et al., 1997), and mean relative performance (MRP) (Osmanzai, 1994). However, the different indices have different levels of precision, making comparisons between genotypes difficult. It is generally presumed that good performance under both irrigated and drought conditions leads to high values of STI, MP, HM, MRP, GMP, YSI and YI and generally low values of
TOL and SSI. To improve sorghum yield and its stability in stress environments, there is a need to identify selection indices able to distinguish high yielding sorghum genotypes in these conditions. However, very limited work has been reported for sorghum from Ethiopia. The study was, therefore, aimed at introgression of drought tolerance genes into adapted varieties through marker-assisted backcrossing and assesses the efficiency of indices to identify drought tolerance in sorghum, so that suitable lines can be recommended for cultivation in drought prone areas of Ethiopia.
MATERIALS AND METHODS
Description of study area
Field experiments were conducted in Rama Kebele of Mereblekhe District in central zone of Tigrai, Ethiopia (Figure 1). The location was selected based on the potential of sorghum grown and availability of irrigation. The site is situated at 14° 23’ 39″ N latitude and 038° 48’ 90″ E longitude. Rama is found at an altitude of 1389 meter above sea level, with average minimum and maximum temperatures ranging from 22 to 38°C, respectively, during the study time (December 2018 to May 2019).

Genetic materials
The parental lines used for this backcrossing program were one donor parent “B35” and eight recurrent parents which are released varieties and known farmers’ cultivars (Tseadachimure and Wediaker [local landraces]; Dekeba, Gambella 1107, Macia, Meko, Melkam, and Teshale [released varieties] (Table 1). The donor parent is known for post-flowering drought tolerant and it has been used as source of tolerant genes to drought by the inter-intra-national sorghum breeding programmes. B35 is a 3-gene dwarf genotype, BC1 derivative of IS12555 accession, a durra from Ethiopian and is known for its stay green (Rosenow et al., 1983; , 2002) with a type-A stay-green-delayed onset of leaf senescence (Thomas and Smart, 1993; Thomas and Howarth, 2000). It is well characterized for its stay green and several research groups (Tuinstra et al., 1997; Crasta et al., 1999; Subudhi et al., 2000; Xu et al., 2000; Sanchez et al., 2002) have identified a number of stay green QTL involving B35. B35 is early maturing, long in stature, has short compact panicle with copious number of infertile branches; purple genotype with small seeds covered by glumes, dry leaf midrib and relatively low yield potential (Srinivas et al., 2009; Kassahun et al., 2010). The recurrent parents are generally high yielding under optimum moisture conditions (MoA, 2018) and popular amongst the farmers but susceptible to terminal drought.










 mean yield of all genotypes under stress conditions,
mean yield of all genotypes under stress conditions,  mean yield of all genotypes in irrigated conditions and SI = Stress intensity.
mean yield of all genotypes in irrigated conditions and SI = Stress intensity.
RESULTS AND DISCUSSION
Yield performance
The analysis of variance for grain yield grown under both moisture regimes indicated the presence of a considerable genotypic variation, indicating differential responses to different environmental conditions, thereby suggesting the possibility of selecting better-performing genotypes under both production environments. Mean grain yields that varied widely in water-limited (1.1 for BC2F3_ETSC_16218 to 4.42 t ha-1 for BC2F3_ETSC_16258) and full-irrigation conditions (2.25 for B35 to 5.71 t ha-1 for Dekeba) were 1.93 and 3.7 t ha-1, respectively (Table 2). This showed that an increase of 47.8 % in yield productivity under the later compared to the former. The grain yield under optimum condition revealed that most of recurrent parents showed highest yield compared to the majority of the developed lines. Among the developed lines with higher yield and statistically similar to the recurrent parents were BC2F3_ETSC_16214, BC2F3_ETSC_16216, BC2F3_ETSC_16251, BC2F3_ETSC_16235, BC2F3_ETSC_16139, BC2F3_ETSC_16258, BC2F3_ETSC_16257, BC2F3_ETSC_16242, and BC2F3_ETSC_16223 indicating the potential of these lines under optimum production environments. On the other hand, the developed near isogenic lines showed highest grain yield under stressed condition. Of the 61 lines, four were the top yielding under stressed conditions; BC2F3_ETSC_16258, BC2F3_ETSC_16216, BC2F3_ETSC_16257, and BC2F3_ETSC_16213 with a yield of 4.42, 3.5, 3.1, and 2.83 t ha-1, respectively. The yield under water-stressed conditions (Ys) had good association with yield obtained under non-stressed conditions (Yp), indicating the possibilities of obtaining potential lines for both moisture regimes. For example, near isogenic lines with a good yield performance under both irrigation conditions were BC2F3_ETSC_16258, BC2F3_ETSC_16216, BC2F3_ETSC_16257, BC2F3_ETSC_16251, and BC2F3_ETSC_16141 (Table 2). The consistence performances of the near isogenic lines in the two contrasting (non-stress vis-à-vis stress) environments represent very nearly the same character, determined nearly by the same set of genes (Falconer, 1989). This may probably have the advantage of the possibilities to forecast the performance of genotypes under one condition on the basis of performance obtained under another and can assist breeders in deciding variety development and allocation of the scarce resources (Keneni, 2007). Therefore, indirect selection for such conditions based on the results of optimum conditions may be efficient (Brennan and Byth, 1979; Rosielle and Hamblin, 1981). However, this needs to be supported by a large data from the multi-location-year experiments as many authors disproved the concept that stipulates cultivars selected under favorable environments also suitable to the unfavorable ones (Ceccarelli and Grando, 1996; Banziger and Edmeades, 1997; Banziger et al., 1997; Banziger and Lafitte, 1997) because it is practically impossible to collect together genes responsible for superior performance in all environments into a single genotype (Annicchiarico, 2002).


Drought tolerance indices
The ANOVA for the quantitative selection indices differed significantly for all indices namely SSI, MRP, MP, HM, GMP, STI, YI, TOL and YSI (Table 2). The mean values of each tolerance indices ranged from the highest 1.61 for BC2F3_ETSC_16235 to the lowest 0.12 for BC2F3_ETSC_16258, 3.48 for BC2F3_ETSC_16258 to 1.19 for B35, 4.5 for BC2F3_ETSC_16258 to 1.7 for B35, 4.52 for BC2F3_ETSC_16258 to 1.47 for B35, 4.52 for BC2F3_ETSC_16235 to 1.58 for B35, 1.72 for BC2F3_ETSC_16258 to 0.18 for BC2F3_ETSC_16215, 2.22 for BC2F3_ETSC_16258 to 0.54 for BC2F3_ETSC_16218, 3.33 for BC2F3_ETSC_16235 to 0.42 for BC2F3_ETSC_16258, and 4.27 for BC2F3_ETSC_16235 to 0.98 for BC2F3_ETSC_16258 in that order. The highest values of SSI and TOL belonged to lines; BC2F3_ETSC_16235, BC2F3_ETSC_16218, BC2F3_ETSC_16238, BC2F3_ETSC_16249, BC2F3_ETSC_16242, BC2F3_ETSC_16217 and BC2F3_ETSC_16139, whereas lower values related to BC2F3_ETSC_16258, BC2F3_ETSC_16229, BC2F3_ETSC_16247, BC2F3_ETSC_16213, BC2F3_ETSC_16252, BC2F3_ETSC_16216, BC2F3_ETSC_16149, BC2F3_ETSC_16239, BC2F3_ETSC_16230, and BC2F3_ETSC_16227. For instance, line BC2F3_ETSC_16235 with both greater SSI and TOL values had grain yield of 4.68 and 1.32 t ha-1 under full-irrigation and water-limited, respectively;, therefore, was identified as highly sensitive to moisture stress after anthesis. In contrast, the lower value of SSI and TOL belonged to BC2F3_ETSC_16258 with grain yield of 4.57 t ha-1 under full-irrigation and 4.42 t ha-1 in water-limitedcondition. Therefore, this line is less sensitive to stress. This means that the greater SSI and TOL values, the greater sensitivity to stress, thus a smaller value of these indices is favored, agreeing with other reports (Rosielle and Hamblin, 1981; Ghasem and Farshadfar, 2015).On the other hand, selection based on TOL with minimum yield reduction under stress condition in comparison with non-stress condition failed to identify the most tolerant genotypes (Farshadfar et al., 2013). Similar to TOL, stress susceptibility index (SSI), genotypes with highest values were considered as genotypes with high drought susceptibility and poor yield stability in both moisture regimes. With regard to yield stability index (YSI) near isogenic lines with higher values were related to BC2F3_ETSC_16258, BC2F3_ETSC_16229, BC2F3_ETSC_16143, BC2F3_ETSC_16216, BC2F3_ETSC_16249, BC2F3_ETSC_16141, BC2F3_ETSC_16247, and BC2F3_ETSC_16221 and were also the most stable under stress and non-stress conditions. The lowest values of SSI and TOL as well as the highest values of YSI indicated that SSI, TOL, and YSI indices were able to identify genotypes with higher yields under drought stress rather than under non-stress conditions.
The tolerance indices MRP, GMP, STI, HM, MP and YI measure the higher stress tolerance and yield potential. Accordingly, the highest and consistent values across all indices belonged to the four near isogenic linesBC2F3_ETSC_16258, BC2F3_ETSC_16216, BC2F3_ETSC_16257, and BC2F3_ETSC_16142 and therefore, they were the most tolerant progenies based on all quantitative indices. These lines were the most tolerant genotypes and also had lower values of SSI and TOL (Table 2). Conversely, the lowest values for all quantitative indices related to B35, BC2F3_ETSC_16215, BC2F3_ETSC_16150, BC2F3_ETSC_16254, BC2F3_ETSC_16238, BC2F3_ETSC_16218, BC2F3_ETSC_16233 and BC2F3_ETSC_16244 and, therefore, some of them were stress sensitive and the other stress tolerant (B35) but with low yield potential under both moisture regimes. Generally, this study showed that quantitative indices (MRP, GMP, STI, HM, MP, and YI) were comparable for identifying superior sorghum genotypes under both environments. Different researches have also used different indices for selecting tolerant genotypes in various crops. For instances, SSI and GMP were preferable in common bean (Ramirez and Kelly, 1998), STI and GMP in maize (Khallili et al., 2004) and mung bean (Fernandez, 1992), durum wheat (Nouri et al., 2011; Mohammadi, 2016), safflower (Majidi et al., 2011; Bahramiet al., 2014), HM, YI, MP, GMP, STI in bread wheat (Khakwani et al., 2011;Dorostkar et al., 2015; Ghasemi and Farshadfar, 2015; Amare et al., 2019), Barley (Nazari and Pakniyat, 2010) and sorghum (Sory et al., 2017) implies that they were useful in identifying lines that yield well under well-watered and also relatively well in water-limited condition.
Interrelationships of the drought tolerance indices
To determine the most desirable drought tolerance criteria, the correlation coefficient between grain yield under the well-watered (Yp), water-limited conditions (Ys), and the quantitative indices of drought tolerance were determined (Table 3). The results of the correlation analysis showed that both positive and negative associations, showing that some of the indices are generally similar and dissimilar in genotypic ranking, respectively. The correlation coefficients of grain yield under non-stressed condition (Yp) showed significant positive correlation with grain yield in the stressed environment (Ys) and all of the selection indices except for SSI and YSI. The significant positive correlations between non-stressed and stressed conditions indicated that genotypes that performed well under non-stress also performed well under stress. No significant correlations were observed between Yp and that of SSI and YSI. In the same manner, grain yield under Ys was significantly and positively correlated with all of the indices except for SSI and TOL which were significant negative correlation (Table 3). A positive correlation between TOL and Yp and the negative correlation between TOL and Ys suggested that selection based on TOL will lead to reduction of yield under well-watered conditions. Among the drought tolerant indices that showed strong positive correlation under both non-stress and stress irrigation include; MRP (r= 0.82; 0.91), MP (r=0.91; 0.83), HM (r=0.71; 0.96), GMP (r=0.81; 92), STI (r=0.76; 0.92) and YI (r=0.52; 1.00), respectively. This indicated that the six indices were comparably effective for selecting and predicting better grain-yielding genotypes under both moisture regimes, corroborating with previous reports (Ezatollah et al., 2012; Farshadfar et al., 2013; Sardouie-Nasab et al., 2015; Darzi-Ramandi et al., 2016). The negative associations of SSI and TOL with grain yield under stress indicated that genotypes with low SSI and TOL values had lower yield differences between non-stress and stress environments (Ceccarelli et al., 1998; Rizza et al., 2004; Mehammadi, 2016).SSI showed significant negative correlation with all selection indices except for TOL that showed significant positive association. Moreover, SSI showed a negative correlation with Ys while no significant correlation was detected between Yp and SSI. Thus, SSI index is suitable for identification of genotypes with low yield and tolerance to drought stress (Kharrazi and Rad, 2011). TOL had significant positive association with MP and significant negative correlation with YI and YSI. TOL was not strongly correlated with indices MRP, GMP, HM, YI, MP and STI. Thus, TOL and SSI ranked differently from the other selection. MRP showed strong significant correlation with MP, HM, GMP, STI, YI and YSI but weak with TOL. Indices of MP, YI, STI, GMP, MRP, and HM showed the existence of strong positive correlation among themselves showing their similarity between these indices for genotypes ranking. According to Farshadfar et al., (2001) most suitable indices for selecting stress-tolerant cultivars is an indices which has a relatively strong correlation with the seed yield under stress and non-stress conditions. Therefore, evaluating correlations between stress tolerance indices and the seed yield in both environments can lead to identification of the most suitable indices. Close correlation between MRP and GMP (r = 1.0) that indicates these two indices are identical in genotypes ranking. YSI had strong and positive correlation with HM, GMP, STI and YI but negatively with SSI and TOL. Likewise, the highest correlation (r = 1.00) was observed between mean grain yield of genotypes under stress (Ys) and yield index (YI). So that consistent correlations were also found between SSI and TOL showing they can be used interchangeably for screening under stress condition. In conclusion, the strong significant positive correlations between HMP, GMP, MP and STI indices showed genotypes with a good performance in both conditions (Yp and Ys) displaying that they are the best indices for identification of superior genotypes agreeing with reports of Mardeh et al. (2006), Golabadi et al. (2006) and Farshadfar et al. (2012).

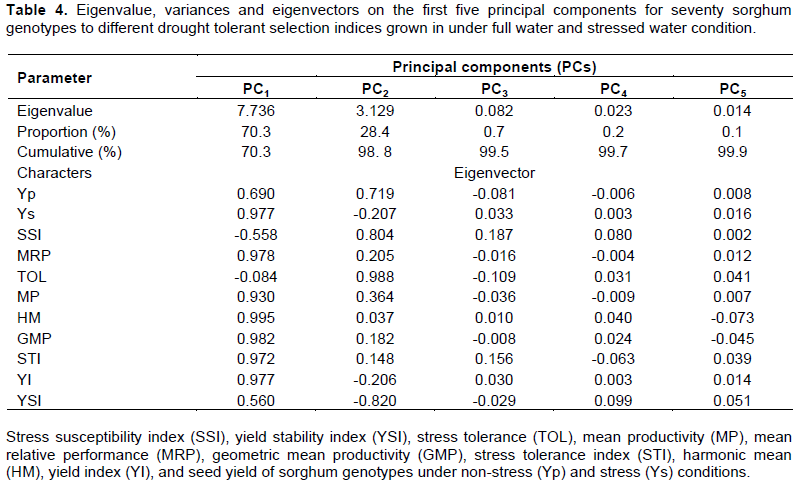
Principal components analysis
Principal components (PC) of the grain yield under water-limited and well-watered conditions as well as drought tolerance indices of the sorghum lines are given in Table 4. The PC analysis was performed to assess the relationships between all attributes to identify superior genotypes under the two-contrasting environments. The results showed that the first five principal components (PC1-PC5) accounted for 99.9% of the entire variation. The first two components grossly explained 98.8% of total variation between the variables (Figure 2). The PC1 alone contributed the largest component score of 70.3% with high positive weight due to grain yield in the stress (Ys) (0.977), MRP (0.978), MP (0.93), HM (0.995), GMP (0.982), STI (0.972), and YI (0.977). Therefore, characters with relatively larger absolute values of eigenvector weights in PC1 had the largest contribution to the differentiation of the genotypes into clusters. It is normally assumed that characters with larger absolute values closer to unity within the first PC influence the clustering more than those with lower absolute values closer to zero (Chahal and Gosal, 2002). The second PC explained 28.4% of the total variation and with high weight corresponding to Yp (0.719), SSI (0.804) and TOL (0.988) due to lower value is preferred for the lower sensitivity to moisture stress and YSI (-0.820); therefore, it was grouped as drought sensitive. This study was in agreement with earlier reports that stated more than 99% of the total variation was explained by the first two principal components (Drikvand et al., 2012; Nouraein et al., 2013; Amare et al., 2019). They also pinpointed the high association of STI, MRP, GMP, HM, MP, and YI with higher grain yield under both conditions. Therefore, selection efforts based on these indices may be more effective. PC1 and PC2 were explained for grain yield potential under both irrigation conditions and stress susceptibility under stressed condition, respectively. This indicates that selecting genotypes with high PC1 and low PC2 is suitable for both moisture regimes (Figure 2). Accordingly genotypes; 4 (BC2F3_ETSC_16141), 17 (BC2F3_ETSC_16213), 20 (BC2F3_ETSC_16216), 52 (BC2F3_ETSC_16248), 61(BC2F3_ETSC_16257) and 62 (BC2F3_ETSC_16258) with high PC1 and low PC2 (low sensitivity and high yield) are likely better genotypes in both environments. These genotypes also showed high values of STI, MP, MRP, YI, MP, GMP and HM as well as low values of SSI and TOL. Whereas, genotypes 5 (BC2F3_ETSC_16142), 18 (BC2F3_ETSC_16214), 55 (BC2F3_ETSC_16251), 63 (Dekeba), 64 (Gambella1107), 65 (Macia), 66 (Meko), and 70 (Wediaker) with both high PC1 and PC2 are suitable in non-stress condition because they are sensitive to terminal drought. On the other side, sorghum genotypes with both low PC1 and PC2 had low sensitivity to stress condition but with low yield potential and can be used in breeding programs for drought tolerance (eg. B35). Conversely, genotypes with low PC1 and high PC2 exhibited inferior yield performance and high sensitivity to end-season drought and therefore their cultivation and incorporating in the breeding programmes may not encouraged. Finally, the two first PCs ascertained that their discrimination and correlation between yield potential and drought sensitively agreeing with earlier reports (Thomas et al., 1995; Kaya et al., 2002; Nazari and Pakniyat, 2010; Nouri et al., 2011; Dorostkar et al., 2015; Ghasemi and Farshadfar, 2015).

Cluster analysis
Cluster analysis based on grain yield under stressed and non-stressed conditions and drought tolerance indices were classified into three clusters (Figure 3). Clustering indices was performed to verify the accuracy of conclusions based on their similarity by average linkage method. Clusters I, II, and III encompassed 64.3, 20 and 15.7% of the genotypes, respectively. The first cluster (C1, n = 45) had the largest number of genotypes and was characterized by high and lowest yield under full-irrigation and water-limited condition, respectively. This cluster also showed lowest values of mean MRP, GMP, MP, STI, HM, YI and YSI, while higher values of SSI and TOL. The cluster constituted those genotypes characterized by overall inferior performances. The second cluster (C2, n = 14) classified as intermediate in mean yield under the two-contrasting moisture regimes and high values of MRP, GMP, MP, STI, HM, YI, and YSI, with lower values of TOL and SSI.

Genotypes in cluster III (C3, n = 11) had high grain yield both under non-stressed (4.52-4.76 t ha-1) and stressed (3.1-4.42 t ha-1) conditions and had the highest value of MRP, GMP, MP, STI, HM, YI and YSI, while lower values of SSI and TOL. This cluster was also superior to grand mean of all other traits averaged over all clusters, indicating that this cluster contained desirable genotypes according to yield obtained from both environments and selection indices. This study is in line with previous reports that stated genotypes can be classified adapted to moisture-stressed and non-stressed conditions using cluster analysis in various crops (Eivazi et al., 2013; Johari-Pireivatlou, 2014; Bahrami et al., 2014; Sory et al., 2017). Generally, this study showed that selection can be improved though MRP, MP, GMP, STI, and HM.
CONCLUSIONS
The results showed significant variations among the developed near isogenic lines, resulting in considerable variation in yield and drought tolerance that could be exploited in sorghum improvement. According to the correlation and principal component analysis, drought tolerance indices MRP, MP, GMP, STI, and HM, and YI are superior indices to identify genotypes that yield well under stressed and optimal conditions. YSI was also found to be more useful indices to discriminate tolerant genotypes that are stable in different conditions and produce high grain yield under stressed conditions. The progenies with high TOL and SSI had high yield only under irrigated conditions and significant yield reduction under stressed conditions.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors appreciate the financial support of Ethiopian Institute of Agricultural Research (EIAR), Agricultural Growth Program-two (AGP-2) and Melkassa Agricultural Research Center (MARC) and for the technical support from Mekhoni Agricultural Research Centre (MkARC) and Axum Agricultural Research Centre (AxARC).
REFERENCES
|
Amare A, Mekbib F, Tadesse W, Tesfaye K (2019). Screening of drought tolerant bread wheat (TriticumaestivumL.) genotypes using yield based drought tolerance indices. Ethiopian Journal of Agricultural Sciences 29(2):1-6. |
|
|
Amelework B, Shimelis H, Tongoona P, Laing M (2015). Physiological mechanisms of drought tolerance in sorghum, genetic basis and breeding methods: a review. African Journal of Agricultural Research 10(31):3029-3040. |
|
|
Annicchiarico P (2002). Genotype x Environment Interaction: Challenges and Opportunities for Plant Breeding and Cultivar Recommendation. FAO Plant Production and Protection Paper No. 174. Food and Agriculture Organization, Rome. P. 174. |
|
|
Araus JL, Cairns JE (2014). Field high-throughput phenotyping: the new crop breeding frontier. Trends in Plant Science 19(1):52-61. |
|
|
Bahrami F, Arzani A, Karimi V (2014). Evaluation of yield-based drought tolerance indices for screening safflower genotypes. Agronomy Journal 106(4):1219-1224. |
|
|
Banziger M, Betran FJ, Lafitte HR (1997). Efficiency of high-nitrogen selection environments for improving maize for low-nitrogen target environments. Crop Science 37:1103-1109. |
|
|
Banziger M, Edmeades GO (1997). Predicted productivity gains from breeding maize under stressed vs. non-stressed conditions. In: Ransom JK, Palmer AFE, Zambezi BT, Mduruma ZO, Waddington SR, Pixley SR, and Jewell DC (Eds.), Maize Productivity Gains through Research and Technology Dissemination: Proceedings of the 5th Eastern and Southern Africa Regional Maize Conference (Arusha, Tanzania, 3-7 June 1996). CIMMYT, Addis Abeba, Ethiopia. pp. 136-140. |
|
|
Banziger M, Lafitte HR (1997). Breeding for N-stressed environments: How useful are N-stressed selection environments and secondary traits? In: Edmeades GO, Banzinger M, Mickelson HR, Pena-Valdivia CB (Eds.), Developing drought and low N-tolerant maize, Proceedings of a Symposium, March 25-29, 1996, CIMMYT, ElBatan, Mexico. Mexico D.F., CIMMYT. pp. 401-404. |
|
|
Bates BC, Kundzewicz ZW, Wu S, Palutikof JP (2008). Climate change and water.Intergovernmental Panel on ClimateChange, Geneva, 210 pp. |
|
|
Blum A (2011). Drought resistance: is it really a complex trait? Functional Plant Biology 38:753-757. |
|
|
Bouslama M, Schapaugh WT (1984). Stress tolerance in soybean.Part 1: evaluation of three screening techniques forheat and drought tolerance. Crop Science 24:933-937. |
|
|
Brennan PS, Byth DE (1979). Genotype and environmental interactions for wheat yields and selection for widely adapted wheat genotypes. Australian Journal of Agricultural Research 30:221-232. |
|
|
Ceccarelli S, Grando S (1996). Importance of specific adaptation in breeding for marginal conditions. In: HailuGebre, Van Leur J (eds.) Barley Research in Ethiopia: Past Work and Future Prospects. Proceedings of the first barley research review workshop. 16-19 October 2003. Addis Ababa: IAT/ICARDA. Addis Ababa, Ethiopia. pp. 34-58. |
|
|
Ceccarelli S, Grando S, Impiglia A (1998). Choice of selection strategy in breeding barley for stress environments. Euphytica 103:307-318 |
|
|
Central Statistical Agency (CSA) (2018). Area and production of majorcrops (private peasant holdings, meher season).Statistical bulletin 586, Addis Ababa, Ethiopia. pp. 1-10. |
|
|
Chahal GS, Gosal SS (2002). Principles and procedures of plant breeding: biometrical and conventional approach. Narosa Publishing House, New Delhi. |
|
|
Charrad M, Ghazzali N, Boiteau V, Niknafs A (2014). NbClust Package for Determining the Best Number of Clusters. R package version 2.0.3, URL |
|
|
Crasta OR, Xu WW, Rosenow DT, Mullet J, Nguyen HT (1999). Mapping of post-flowering drought resistance traits in grain sorghum: association between QTLs influencing premature senescence and maturity. Molecular and General Genetics MGG 262(3):579-88. |
|
|
Darzi-Ramandi H, Najafi-Zarini H, Razavi K, Kazemitabar S (2016). Screening Iranian bread wheat lines under different water regimes using yield based drought tolerance indices. SABRAO Journal of Breeding and Genetics 48(4):491-503. |
|
|
Dorostkar S, Dadkhodaie A, Heidari B (2015). Evaluation of grain yield indices in hexaploid wheat genotypes in response to drought stress. Archives of Agronomy and Soil Science 61(3):397-413. |
|
|
Drikvand R, Doosty B, Hosseinpour T (2012). Response of rainfed wheat genotypes to drought stress using drought tolerance indices. Journal of Agricultural Science (Toronto) 4(7):126-131. |
|
|
Eivazi AR, Mohammadi S, Rezaei M, Ashori S, Pour FH (2013). Effective selection criteria for assessing drought tolerance indices in barley (Hordeumvulgare L.) accessions. International Journal of Agronomy and Plant Production 4(4):813-821. |
|
|
Falconer DS (1989). Introduction to Quantitative Genetics. 3rd ed. Longman, London, England |
|
|
Food and Agriculture Organization (FAO) (2017). Food and Agriculture Organization of the United Nations, Rome, Italy. |
|
|
Farshadfar E, Moradi Z, Elyasi P, Jamshidi B, Chaghakabodi R (2012). Effective selection criteria for screening drought tolerant landraces of bread wheat (Triticumaestivum L.). Annals of Biological Research 3(5):2507-2516. |
|
|
Farshadfar E, Poursiahbidi MM, Safavi SM (2013). Assessment of drought tolerance in land races of bread wheat based on resistance/tolerance indices. International Journal of Advanced Biological and Biomedical Research 1(2):143-58. |
|
|
Farshadfar E, Zamani M, Motallebi M, Imamjomeh A (2001). Selection for drought resistance in chickpea lines. Iranian Journal of Agricultural Sciences 32(4):65-77. |
|
|
Fernandez GCJ (1992). Effective selection criteria for assessing plant stress tolerance. pp. 257-270. In: Kuo CG (ed.) Adaptation of Food Crops to Temperature and Water Stress: Proceedings of an International Symposium, Tainan, Taiwan. |
|
|
Fischer RA, Maurer R (1978). Drought resistance in spring wheat cultivars. I. Grain yield responses. Australian Journal of Agricultural Research 29(5):897-912. |
|
|
Gavuzzi P, Rizza F, Palumbo M, Campanile RG, Ricciardi GL, Borghi B (1997). Evaluation of field and laboratory predictors of drought and heat tolerance in winter cereals. Canadian Journal of Plant Science 77(4):523-531. |
|
|
Ghasemi M, Farshadfar E (2015). Screening drought tolerant genotypes in wheat using multivariate and stress tolerance score methods. International Journal of Biosciences 6:326-333. |
|
|
Golabadi M, Arzani AS, Maibody SM (2006). Assessment of drought tolerance in segregating populations in durum wheat. African Journal of Agricultural Research 1(5):162-171. |
|
|
Harris K, Subudhi PK, Borrell A, Jordan D, Rosenow D, Nguyen H, Klein P, Klein R, Mullet J (2007). Sorghum stay-green QTL individually reduce post-flowering drought-induced leaf senescence. Journal of Experimental Botany 58(2):327-338. |
|
|
Hossain AB, Sears RG, Cox TS, Paulsen GM (1990). Desiccation tolerance and its relationship to assimilate partitioning in winter wheat. Crop Science 30(3):622-627. |
|
|
Johari-Pireivatlou M (2014). Selection for drought tolerance in bread wheat genotypes with drought tolerance and susceptibility indices. Journal of Applied Environmental and Biological Sciences 4:128-133. |
|
|
Kassahun B, Bidinger F, Hash C, Kuruvinashetti M (2010). Stay-green expression in early generation sorghum [Sorghum bicolor (L.) Moench] QTL introgression lines. Euphytica 172:351-362. |
|
|
Kaya Y, Palta C, Taner S (2002). Additive main effects and multiplicative interactions analysis of yield performances in bread wheat genotypes across environments. Turkish Journal of Agriculture and Forestry 26(5):275-279. |
|
|
Keneni G (2007). Concerns on mismatches between environments of selection and production of crop varieties in Ethiopia. East African Journal of Sciences 1(2):93-103. |
|
|
Khakwani AA, Dennett MD, Munir M (2011). Drought tolerance screening of wheat varieties by inducing water stress conditions. Songklanakarin Journal of Science and Technology 33(2):135-142. |
|
|
Khallili M, Kazemi A, Moghaddam A, Shakiba M (2004). Evaluation of drought toleranceindices at different growth stages of late-maturing corn genotypes. In: The 8th Iranian Congress of Crop Sciences and Breeding. 25-27 August 2004, Rasht, Iran. Rasht: University Press. pp. 298-298. |
|
|
Kharrazi MA, Rad MR (2011). Evaluation of sorghum genotypes under drought stress conditions using some stress tolerance indices. African Journal of Biotechnology 10(61):13086-13089. |
|
|
Kumar AA (2016). Botany, Taxonomy and Breeding. In: Rakshit S, Wang Y-H (eds.), The Sorghum Genome, Compendium of Plant Genomes, |
|
|
Madhusudhana R (2015). Application of DNA Markers for Genetic Improvement. In: Madhusudhana R, Rajendrakumar P, Patil JV (eds). Sorghum Molecular Breeding. Springer pp. 71-99. |
|
|
Majidi MM, Tavakoli V, Mirlohi A, Sabzalian MR (2011). Wild safflower species ('Carthamusoxyacanthus'Bieb.): A possible source of drought tolerance for arid environments. Australian Journal of Crop Science 5(8):1055-1063. |
|
|
Mardeh AS, Ahmadi A, Poustini K, Mohammadi V (2006). Evaluation of drought resistance indices under various environmental conditions. Field Crops Research 98(2-3):222-229. |
|
|
Mera GA (2018). Drought and its impacts in Ethiopia. Weather and Climate Extremes 22: 24-35. |
|
|
Ministry of Agriculture (MoA) (2018). Plant Variety release, protection and seed quality control directorate. Crop variety register, issue No. 21, Addis Ababa, Ethiopia pp. 68-77. |
|
|
Mohammadi R (2016). Efficiency of yield-based drought tolerance indices to identify tolerant genotypes in durum wheat. Euphytica 211(1):71-89. |
|
|
Nazari L, Pakniyat H (2010). Assessment of drought tolerance in barley genotypes. Journal of Applied Sciences 10(2):151-156. |
|
|
Nouraein M, Mohammadi SA, Aharizad S, Moghaddam M, Sadeghzadeh B (2013). Evaluation of drought tolerance indices in wheat recombinant inbred line population. Annals of Biological Research 4(3):113-122. |
|
|
Nouri A, Etminan A, Teixeira da Silva JA, Mohammadi R (2011). Assessment of yield, yield-related traits and drought tolerance of durum wheat genotypes (Triticumturjidum var. durumDesf.). Australian Journal of Crop Science 5(1):8-16. |
|
|
Osmanzai M (1994). A screening method for productivity in moisture deficit environments.Sorghum Improvement Conference of North America, USA; University of Georgia, USA; International Crops Research Institute for the Semi-Arid Tropics, Patacheru 502:324, Andhra Pradesh, India. |
|
|
Patterson HD, Thompson R (1971). Recovery of inter-block information when block sizes are unequal.Biometrika, 58:545-554. |
|
|
R Core Team (2019). R: a Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/. |
|
|
Raman A, Verulkar S, Mandal N, Variar M, Shukla V, Dwivedi J, Singh B, Singh O, Swain P, Mall A, Robin S (2012). Drought yield index to select high yielding rice lines under different drought stress severities. Rice 5:1-12. |
|
|
Ramirez P, Kelly JD (1998). Traits related to drought resistance in common bean. Euphytica, 9: 127-136. |
|
|
Reddy NR, Madhusudhana R, Murali MS, Seetharama N, Jagannatha VP (2014). Detection and validation of stay-green QTL in post-rainy sorghum involving widely adapted cultivar, M35-1 and a popular stay-green genotype B35. BMC Genomics 15(909):1471-2164. |
|
|
Reddy PS (2017). Sorghum, Sorghum bicolor (L.) Moench. In: Patil JV (Ed.), Millets and Sorghum: Biology and Genetic Improvement. pp. 1-32. |
|
|
Ribaut JM, Bänziger M, Betran J, Jiang C, Edmeades GO (2002). Breeding: Drought Tolerance Improvement in Tropical Maize. Quantitative Genetics, Genomics, and Plant Breeding pp. 85-99. |
|
|
Rizza F, Badeck FW, Cattivelli L, Lidestri O, Di Fonzo N, Stanca AM (2004). Use of a water stress index to identify barley genotypes adapted to rainfed and irrigated conditions. Crop Science 44(6):2127-2137. |
|
|
Rosenow DT, Clark LE, Dahlberg JA, Frederiksen RA, Odvody GN, Peterson GC, Miller FR, Woodfin CA, Schaefer K, Collins SD, Jones JW (2002). Release of four A/B sorghum parental lines ATx642 through ATx645. International Sorghum and Millets Newsletter 43:24- 30. |
|
|
Rosenow DT, Quisenberry JE, Wendt CW, Clark LE (1983). Drought tolerant sorghum and cotton germplasm. In: Developments in agricultural and managed forest ecology Elsevier 12: 207-222. |
|
|
Rosielle AA, Hamblin J (1981). Theoretical aspects of selection for grain yield in stress and non-stress environments. Crop Science 21:943-946. |
|
|
Sabadin PK, Malosetti M, Boer MP, Tardin FD, Santos FG, Guimaraes CT, Gomide RL, Andrade CL, Albuquerque PE, Caniato FF, Mollinari M (2012). Studying the genetic basis of drought tolerance in sorghum by managed stress trials and adjustments for phenological and plant height differences. Theoretical and Applied Genetics 124(8):1389-1402. |
|
|
Sanchez AC, Subudhi PK, Rosenow DT, Nguyen HT (2002). Mapping QTLs associated with drought resistance in sorghum (Sorghum bicolorL. Moench). Plant Molecular Biology 48(5-6):713-726. |
|
|
Sardouie-Nasab S, Mohammadi-Nejad G, Nakhoda B (2015). Field screening of salinity tolerance in Iranian bread wheat lines. Crop Science 54(4):1489-1496. |
|
|
Schneider KA, Rosalesâ€Serna R, Ibarraâ€Perez F, Cazaresâ€Enriquez B, Acostaâ€Gallegos JA, Ramirezâ€Vallejo P, Wassimi N, Kelly JD (1997). Improving common bean performance under drought stress. Crop Science 37(1):43-50. |
|
|
Sharma KK, Lavanya M (2002). Recent developments in transgenics for abiotic stress in legumes of the semi-arid tropics. JIRCAS Working Report No. 23(23):61-73. |
|
|
Singh BD (2002). Plant Breeding: Principles and Methods. Kalyani Publishers, New Delhi-Ludhiana. |
|
|
Sory S, Gaoussou DA, Mory CM, Niaba T, Gracen V, Eric D (2017). Genetic analysis of various traits of hybrids sorghum (Sorghum bicolor(L) Moench), correlated with drought tolerance. Journal of Plant Biology and Soil Health 4(1):1-9. |
|
|
Srinivas G, Satish K, Madhusudhana R, Reddy NR, Mohan M, Seetharama SN (2009). Identification of quantitative trait loci for agronomically important traits and their association with genic-microsatellite markers in sorghum. Theoretical and Applied Genetics 118:1439-1454. |
|
|
Subudhi PK, Rosenow DT, Nguyen HT (2000). Quantitative trait loci for the stay green trait in sorghum (Sorghum bicolor L. Moench): consistency across genetic backgrounds and environments. Theoretical and Applied Genetics 101(5-6):733-741. |
|
|
Teshome A, Zhang J (2019). Increase of Extreme Drought over Ethiopia under Climate Warming. Advances in Meteorology 2019:1-18. |
|
|
Thomas H, Dalton SJ, Evans C, Chorlton KH, Thomas ID (1995). Evaluation drought resistance in germplasm of meadow fescue. Euphytica 92:401-411. |
|
|
Thomas H, Howarth CJ (2000). Five ways to stay green. Journal of Experimental Botany 51:29-337. |
|
|
Thomas H, Smart CM (1993). Crops that stay green. Annals Applied Biology 123:193-233. |
|
|
Tuinstra MR, Grote EM, Goldsbrough PB, Ejeta G (1997). Genetic analysis of post-flowering drought tolerance and components of grain development in Sorghum bicolor (L.) Moench. Molecular Breeding 3:439-448. |
|
|
Visarada KBRS, Aruna C (2019). Sorghum: A Bundle of Opportunities in the 21st Century. In: Aruna C, Visarada KBRS, Venkatesh Bhat B, Tonapi VA (eds). Breeding Sorghum for diverse end uses pp. 1-14. |
|
|
Wagaw K (2019). Review on Mechanisms of Drought Tolerance in Sorghum (Sorghum bicolor (L.) Moench) Basis and Breeding Methods. Academic Research Journal of Agricultural Science and Research 7(2):87-99. |
|
|
Wassmann R, Jagadish SV, Sumfleth K, Pathak H, Howell G, Ismail A, Serraj R, Redona E, Singh RK, Heuer S (2009). Regional vulnerability of climate change impacts on Asian rice production and scope for adaptation. Advances in Agronomy 102:91-133. |
|
|
Xu W, Subudhi PK, Crasta OR, Rosenow DT, Mullet JE, Nguyen HT (2000). Molecular mapping of QTLs conferring stay-green in grain sorghum (Sorghum bicolor L. Moench). Genome 43(3):461-469. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


