Full Length Research Paper

ABSTRACT
Okra has significant area under cultivation in Africa and Asia, despite it has been considered as a minor crop and not much attention has been given to okra improvement in international research program. Niger produced during 2008, 2794 t of okra in 5288 ha with mean productivity of 0.39 t.ha-1. This poor average productivity is far below than the mean productivity of East Africa (6.2 t.ha-1) and North (8.8 t.ha-1). Non-availability of varieties resistant to local biotic stresses is one of the major reasons for such a low productivity. Root-knot nematode disease is one of the constraints of commercial okra cultivation in region and cause of poor productivity. Durable host plant resistance/tolerance is considered as better option than the management tactic solely based on insecticide spray alone. The major objective of this study initited by AVRDC is to screening okra germplasm against root knot nematode in order to identify resistant lines. A total of 48 lines belonging to two species (A. esculentus and A. caillei) originated from different countries were screened following the protocol developed during this study. The results showed that after five weeks of inoculation, infestation of nematode is very fatal for susceptible okra genotypes. Cluster analysis using various parameters revealed that groups GI, GV and GII represented lines which are showing less gall index and poor reproduction rate of nematode, whereas GIII and GIV represented lines with more gall index with higher reproduction rate. The lines showing reaction towards resistance can be used in the breeding program.
Key words: Abelmoschus species, okra germplasm, Meloidogyne species, root knot, nematode, screening.
INTRODUCTION
Okra belongs to the family Malvaceae. This family is represented in Niger by several cultivated species such as sorrel (Hibiscus sabdariffa L) cultivated for its calyx and seeds, and okra (Abelmoschus esculentus L.) for its fruits. Okra is characterized by diversity in the shape and color of fruits and stems (Ouédraogo, 2016; Idi-Saidou et al., 2019). It is a vegetable that is found fresh, and in slices (dried or powdered), in all markets during the winter period and the dry season. This is due to its high mucilage content, high market value and vital importance in feeding urban and rural populations (In Situ Conservation Project, INERA/BF, 2001). There are some okra varieties that are recommended to facilitate or enrich the nutrition of the sick. Okra is of considerable economic importance for women and plays a vital role in the nutritional balance of rural populations. The fruit is indeed rich in carbohydrates (7 to 8% of the dry matter) present in the form of mucilage. It is fairly low in fiber but high in protein for a vegetable fruit (1.8% of the dry matter), aspartic acid and arginine account for 10% of amino acids. Despite average vitamin A content (300 IU), the contents of thiamine, riboflavin, and ascorbic acid (vitamin C) are good. Daily consumption of 100 g of fresh okra would provide about 20% of calcium requirements, 15% of iron requirements and 50% of vitamin C requirements (Hamon, 1988; Agbo et al., 2008; Sorapong, 2012); it contains little calcium (90 mg/100 g), phosphorus (56 mg) and magnesium (43 mg/100 g), and very little potassium. According to Hamon (1988), the most important work was done in India and the Southern United States. Importance of the pigmentation characteristics of the various parts of the plant and fruit forms, leaf types, and degree of pubescence (Siemonsma and Hamon, 2004) argues that the study of the genetic variability of the genus Abelmoschus have not really been addressed. Very little information is available on the genetic variability of the cultivated species A. esculentus in general, and in particular in West Africa (Ouédraogo, 2009; Ouédraogo, 2016). Africa in the West is based on the agro-morphological characterization of certain parameters such as plant density, productivity (Fondio et al., 2003, 2011) or on certain interactions between the genotype and the environment (Alake and Ariyo, 2012). None of these studies are carried out under similar agro-ecological conditions of our sites. Moreover, the physiological aspects of this plant are nowhere discussed.
Okra culture remains a predominantly female activity in Niger. From sowing to harvesting, the fruits, leaves, and seeds of okra are handled by the feminine people, who are particularly fond of it for its many uses: sauce, salad, donuts and nutrition of the sick. Okra is also a reliable source of income for women who farm more than 39, 25, 26, 4.5, 3.2 and 0.4% of the area occupied by this crop, respectively in the six regions Tahoua, Tillaberi, Maradi, Zinder, urban community of Niamey and Agadez, located in five different agro-ecological zones of Niger: The Saharan zone (with average annual rainfall is less than 200 mm), the Sahara-Sahelian zone (with an average annual rainfall of between 200 and 300 mm), the Sahelian zone (with an average annual rainfall of between 300 and 400), the Sahelo-Sudan zone (with an average rainfall of between 400 and 600 mm) and the Sudanese zone (with an average annual rainfall of over 600 mm of water per year) (DS/MDA, 2008: Year Book of Statistics Edition 2008: 128).
Young fresh fruits of okra are more appreciated. In fact, when they are 3 to 5 cm long, they are harvested and sold in the market square at a rate of four to five fruits per 100 to 150 CFA francs (In Situ Conservation Project, INERA/BF, 2001). These already high costs are often increased by 50 to 60% during the dry periods of the year.
Despite its multiple uses, its proven nutritional value and its financial value, okra is cultivated on very small surfaces, near the huts and often in the dumps. The cultivation of improved varieties of okra in Niger is not yet reported. These are often local ecotypes that are observed in Nigerian farmers.
Idi-Saidou et al. (2019) is of the view that the ecotype is a local breed (ecological race) whose genotypes are adapted to a specific habitat limited by natural selection in the local environment. These ecotypes cultivated by peasants would probably be of more populations than pure lines. In addition to this, is the drought, the diseases and the attacks of the insect pests which cause a significant reduction of its production (Dabiré-Binso et al., 2009; Sanwal et al., 2014, 2016; Ratnadass et al., 2014; Kumar and Chowdhury, 2019), thus bringing to play the last roles in the country's agricultural research programs, despite the existence of a variety of ecotypes that can be used in the creation of short-cycle varieties or resistant to certain abiotic or biotic constraints. The low rate of existence of improved varieties argues in favor of a participatory selection strategy that takes into account the selection criteria of farmers, integrating the tolerance or resistance of the genotype to the drastic conditions of the environment such as root attack from okra by nematodes (case of genus Meloidogyne) to critical periods of the realization of its genetic potential.
Since the beginning of the 1980s, there has been a noticeable increase in interest in market gardening in Niger. This phenomenon is partly explained by the efforts made by the various governments to control the effects of chronic droughts exacerbated by the degradation of natural resources (land, water, plant cover, etc.) and the extensive nature of our agriculture. Traditionally, farmers grow some vegetables (okra, sorrel) in combination with food crops (millet, sorghum). Production was then exclusively consumed. Today vegetable crops have become real cash crops (onion, pepper). The areas assigned to them are estimated at 48000 ha (DS/MDA, 2008), for an annual production of more than 700 000 t. Part of this production is sold in local markets and the surplus is exported to neighboring countries (Ivory Coast, Nigeria, Togo, and Ghana), thus providing a significant foreign exchange to our country. The value of this production was 45 billion in 1999 (FAO, 2004). Despite the importance of crops in the national economy, there is rather limited attention to the phytosanitary problems that affect them in general and to parasitic nematodes in particular (Haougui, 1999; Haougui and Kollo, 2008). The latter, however, are formidable enemies of market gardeners because they are a limiting factor in the production of their crops.
The primary objective of our work is to determine the resistant varieties of okra following the artificial infestation of the nematodes of the genus Meloidogyne through the evaluation of the elements of the yields, the nematode multiplication rate and the gall index at the root level of these plants. This makes it possible to evaluate the level of the anatomical-morphological and physiological reactions of the different morphotypes under the conditions of the attack of these varieties of okra by these nematodes, and to retain the best genotypes for an improvement program.
MATERIALS AND METHODS
Plant
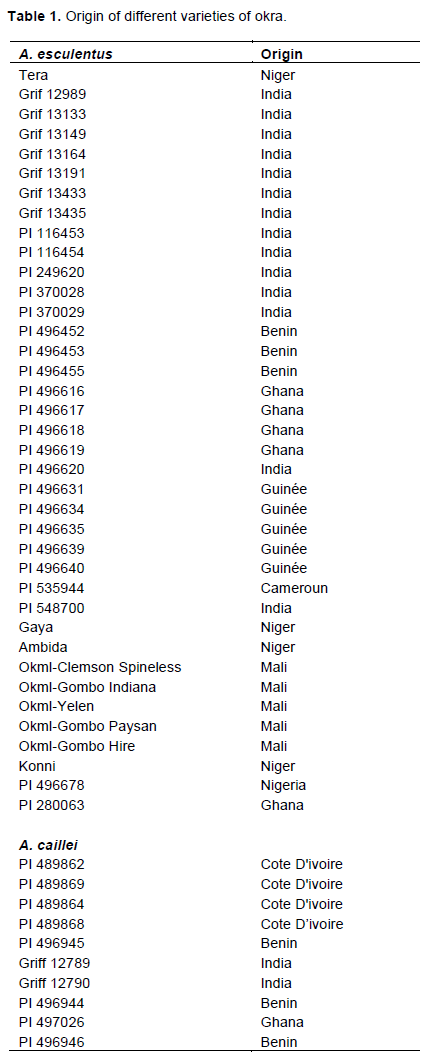
The plant material used in this study consists of 48 varieties of okra (37 of A. esculentus and 11 of Abelmoschus caillei). These varieties of various origins are collected by the AVRDC program based on ICRISAT/NIGER (Table 1).
The root-knot nematodes
The root-knot nematodes were extracted juveniles of the second stage (J2) of Meloidogyne spp. from a tomato farm CV Xina at INRAN's Department of Rural Economic Research.
Germination test
The germination test was carried out by placing 20 seeds of each variety that had been used as plant material, in Petri dishes containing a filter paper and then watered with a small blade of water. The sprouts were then counted every two days for two weeks.
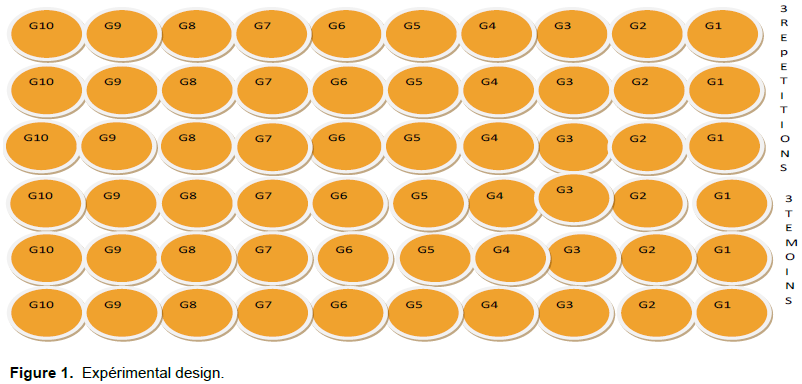
Experimental design and culture conditions
The test on the screening of okra varieties was conducted at the ICRISAT Sahel Center in Sadoré (located in southwestern Niger, 45 km south of the capital Niamey) during the winter 2009 campaign carried out in the plot of the culture of the plants of the various species intended to be transplanted in the fields. The experimental setup (Figure 1) was installed under a shed enclosed by a transparent cover resembling an impregnated net. The 288 pots of culture were placed on metal tables under the shed. Among the six (6) pots selected for each genotype, three pots received the inoculum while the other three pots are considered as witnesses to the test. The different morphological and nematological parameters of these different varieties were measured 5 weeks after inoculation. This allowed appreciation in each variety of the degree of sensitivity towards these plant parasitic diseases. Climatic data were collected using equipment placed under the hangar during the experiment. All varieties used for the trial were sown on 03/11/2009 at the rate of 4 seeds per pot. Before sowing, the seeds of each variety were put in a cup full of water. Only seeds that have settled at the bottom of the cup were used for sowing. No preliminary treatment was carried out on these seeds before their cultivation. Weeds are the main factors limiting the production of okra. To avoid competition for nutrients, water, and light, weeding was done by hand on the 28th day after sowing. A single plant per pot according to the sowing method was carried out at the time of weeding. Its goal is to reduce competition between plants and to allow better use of available resources. The collected plants were used to replace the missing pots within the same variety.
Method of extracting the infesting stage (J2) of Meloidogyne in roots
Knotted roots from tomatoes infested with INRAN gall nematodes were sent to the Nematology Laboratory of the AGRHYMET Center for the Extraction of Juveniles of the 2nd stage of Meloidogyne spp. Here, Seinhorst (1950) method involves washing the infested roots to rid them of the soil used. They were then cut into small fragments of 1 to 1.5 cm and placed in a sieve placed on a funnel whose end is extended by a flexible hose. Everything is placed in a fog chamber for a week. J2 harvesting was done on the 2nd and 3rd day after placement of the gall roots in the fog chamber. Photos 1 and 2 show some materials used for the extraction of nematodes.
Method of extraction of nematodes in the soil
After the seed was removed, the soil was washed by the bucket method to extract the nematodes. This method involves diluting the soil sample in water and then stirring vigorously with the hand. After 2 to 3 min of rest, the liquid suspension is passed through a 25-μm diameter sieve which prevents the passage of nematodes and other debris. This operation is repeated three times in order to collect the maximum number of nematodes. The suspensions collected are then poured onto kleenex papers placed in plates containing water. Thanks to their mobility, the nematodes cross the paper kleenex and are found at the bottom of the plates. After 48 h of active passage, the nematodes are collected on a 25 μm diameter sieve and the contents are put into test tubes.
The enumeration of nematodes collected from soil samples or roots was done using a binocular magnifying glass. Only species (Meloidogyne javanica, Meloidogyne arenaria, and Meloidogyne incognita acrita) belonging to the genus Meloidogyne were taken into account.
Inoculation method and conduction of the test
For application of the inoculum, three holes were made with the tip of the pencil all around the two weeks old okra plant using a graduated pipette. 1 ml of the inoculum containing 100 larvae of the 2nd stages (J2) was taken and distributed in the three holes which are immediately closed with the fingers. This operation was done in three of the six pots used for each variety tested. After inoculation, the plants were watered twice a day in the amount of 100 to 150 ml per culture pot for five weeks. For fertilization, each pot received 1 g of NPK 19.19.19 fertilizer.
Methods of data collection
Parameters that were measured during the test include agronomic and nematological parameters.
Agronomic parameters
Measurement of the agronomic parameters involved the following:
i) Counting the number of seeds that sprouted in each pot of culture to appreciate the percentage of emergence thereof.
ii) Taking into consideration the number of days before appearance of flowering to estimate the precocity and lateness of the varieties resulting from the treatment. This parameter corresponds to the number of days that elapsed between sowing and appearance of the first flower.
iii) Bearing in mind the height of the seedlings that is measured from the neck to the top of the terminal bud (this measurement was made 5 weeks after inoculation).
iv) Taking note of aboveground biomass (fresh weight and dry weight of the plant) and root biomass (fresh weight and dry weight of the root) were 5 weeks after nematode inoculation.
Nematological parameters
This involves evaluation of the gall index using the Zeck (1971) scale which consists of the degree of infestation of nematodes (Table 2)

The final population of juveniles (J2) in each pot was tested to estimate nematode reproduction rate. This rate is calculated using the following formula:
Rate of nematode reproduction = Pf / Pi
where Pf: Nematode final population and Pi: Initial nematode population.
Data analysis
For the analysis of data, three softwares were used. The SAS (Version 9.1) which allowed us to calculate the averages of the measured parameters, the Minitab14 and SAS 9.1 for the analysis of variance and the multivariate analysis from the averages of the various parameters obtained. The different analyses made are: principal component analysis (PCA) to observe the correlations between the different traits from treatments 1 and 2; and the hierarchical ascending classification (CAH) from the coordinates of individuals provided by the ACP. The relative contributions of the axes and the explanatory variables were used to select axes 1 and 2. Groups of similar individuals were constituted by truncation of the hierarchical tree by maximizing the inter-class variance. For the interpretations, we used the GLEGLE Booklets (Glegle and Kokode, 2004).
RESULTS
Germination test in a Petri dish
Okra seeds started to germinate two days after being introduced into the Petri dish. The germination test showed that the percentage of germination varies from 25 to 100% for the varieties of the species A. esculentus, whereas all the varieties of the species A. caillei gave 100% of germination. All seeds sprouted a week after being canned in Petri dish. Varieties with a low germination percentage are from Benin (PI496955), Guinea (PI496639), Mali (Okml Gombo Hire) and India (Grif12790).
Enumeration of juvenile (J2) of Meloidogyne contained in the inoculum
After nematode extraction from the galled roots, we had to count a total of 30000-second instar larvae (J2) in 300 ml of the inoculum, that is to say, we had inoculation 100 J2 larvae of Meloidogyne spp. in 1 ml of the inoculum.
Varieties performance analysis
The analysis of variance of the parameters studied is presented in Table 3.
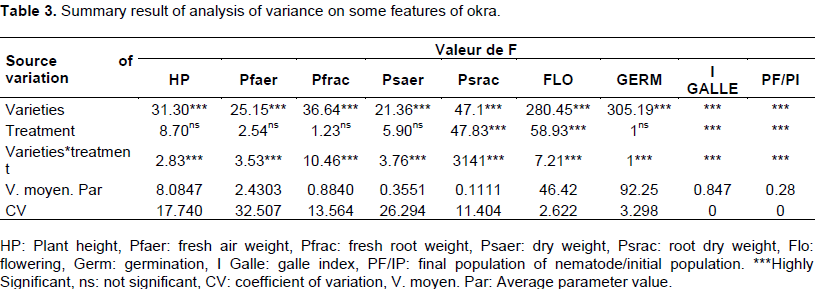
The results of the analysis of variance show that at the variety level, the difference is very highly significant for all the parameters studied as indicated by the values of F (Table 3). This analysis also shows that at the level of the interaction between varieties and treatment, the difference is very highly significant whereas for the treatment, the difference is significant only at the level of the parameters such as root dry weight, flowering time, and flowering time, gall index, and the rate of multiplication of nematode populations. Measured parameters such as plant height, aerial weight, aerial dry weight, and fresh root weight did not show a significant difference in the source of variation used for treatment (Table 3).
It can, therefore, be concluded that there is a great variability within the varieties for the parameters studied. This variability is also important in the interaction between varieties and treatment for the same parameters. On the other hand, at the level of the treatment, the variability was important only within the four studied parameters: root dry weight, flowering time, gall index and nematode multiplication rate (Table 3); which proves the very highly significant differences observed for these parameters.
Analysis of relationships between respective characters
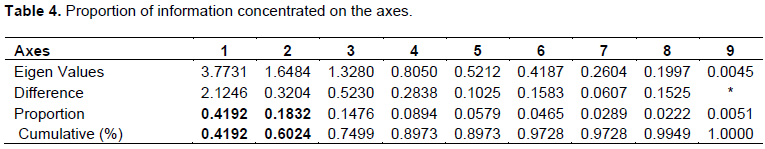
The results of the principal component analysis carried out on the basis of the table of means of parameters (Appendix 1) obtained for the screening of the different varieties of okra reveal that the first two axes (Table 4) account for 60.24% of the information on the characteristics of the varieties. These two axes are therefore retained to explain the relationship between the biological characteristics of the different varieties of okra under the effect of artificial infestation by nematodes of the genus Meloidogyne. To better understand the relationship between the biological characteristics of different varieties of okra under the effect of nematological attack, it was necessary to see the correlations between the studied parameters and the principal components given by the PCA as indicated in Table 5.
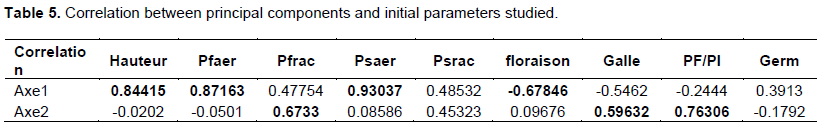
The projection of these different parameters in the axis system defined by the first two main components (Figure 2) shows that for Axis 1 (Explaining 41.92% of information), the highest yield of plant height, fresh aerial weight, and aerial dry weight of okra varieties, are associated with the lowest values of flowering time, gall index, and also the nematode multiplication rate. Thus, this axis is defined as the axis of the resistant varieties with respect to the attack of the inoculated nematodes.
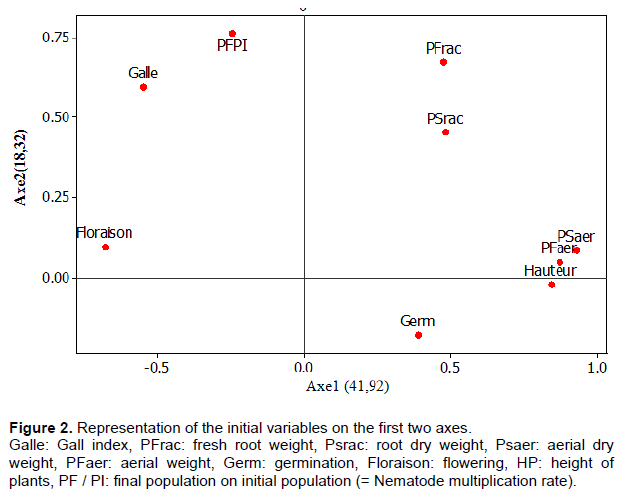
On axis 2 (Explaining 18.32% of information), we note that the highest values of the gall index, the rate of multiplication of nematode populations and fresh root weight are associated with low values of plant height, fresh weight and aerial dry weight, and germination. This axis is defined as the axis of varieties susceptible to nematodes of the genus Meloidogyne. Figure 2 shows the association of these parameters to the different axes in the same way as they were indicated in Table 5.
On the basis of these analyzes, it can be said that, on one hand, the varieties of okra with high values of height, fresh weight, and aerial dry weight, are those with the lowest nematode infestation, indicated by the highest small value of the gall index (0 to 3) (Table 1 of Annex 1), but also by the lowest nematode multiplication rate (PF / PI <1). These varieties, which are both early, are therefore varieties resistant to the phytoparasitic nematodes genus Meloidogyne which have been observed after five weeks of inoculation.
On the other hand, the susceptible varieties were those which showed the highest values of the gall index (5 to 7; Annex 1), the nematode multiplication rate (PF / PI> 1), and fresh root weight. These varieties are those that, by their nature, record low yields of plant height, fresh weight, and dry weight. The same varieties which had been declared to be susceptible showed before the artificial infestation of the nematodes, a bad percentage of germination (25%), which stipulates their weaknesses vis-a-vis the environmental parameters.
Structure analysis of the varieties
Varieties grouping
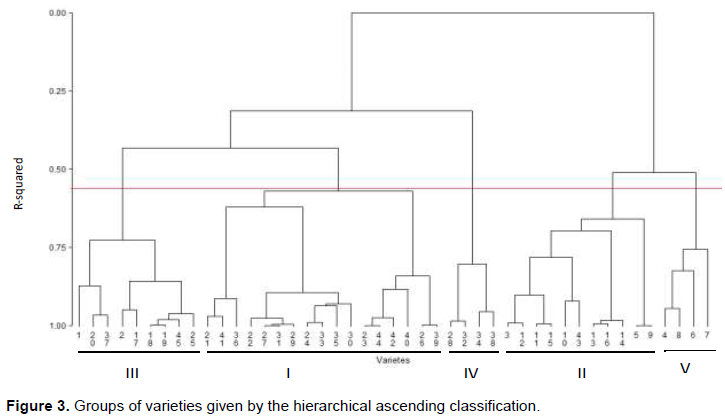
The hierarchical ascending classification (CAH) made from the coordinates of the individuals provided by the ACP gives five groups which consist of the following ways:
Group I consists of varieties 27, 31, 23, 44, 26, 39, 29, 24, 33, 22, 42, 21, 41, 35, 30, 36 and 40. Of these varieties, up to thirteen (13) (21, 22, 31, 35, 23, 26, 41, 24, 27, 39, 33, 29 and 30) belong to the species A. esculentus. The other varieties of the group (42, 44, 41 and 40) belong to the species A. caillei.
Group II comprises varieties 5, 9, 11, 15, 13, 16, 3, 12, 14, 10 and 43. Among these, only variety 43 belongs to the species A. caillei while the others all belong to the species A. esculentus.
Group III is composed of varieties 18, 19, 45, 20, 37, 25, 2, 17, 1 of which only three of them, namely 37, 19 and 17 belong to the species A. caillei while the other varieties belong to the species A. esculentus.
Group V contains varieties 4, 8, 6 and 7, and all belonging to the species A. esculentus.
The total number of varieties at the level of the different groups are respectively 17, 11, 9, 4 and 4 for Groups I, II, III, IV, and V. The average performances of the varieties of each group, characterized by the different studied parameters are shown in Appendix 2. Figure 3 shows the different groups obtained by the hierarchical ascending classification (CAH).
Groups characterization
The projection of these different groups of okra varieties into the system of axes defined by the first two main components (Figure 4) gives us a grouping of entries that cover the groups determined by the CAH.
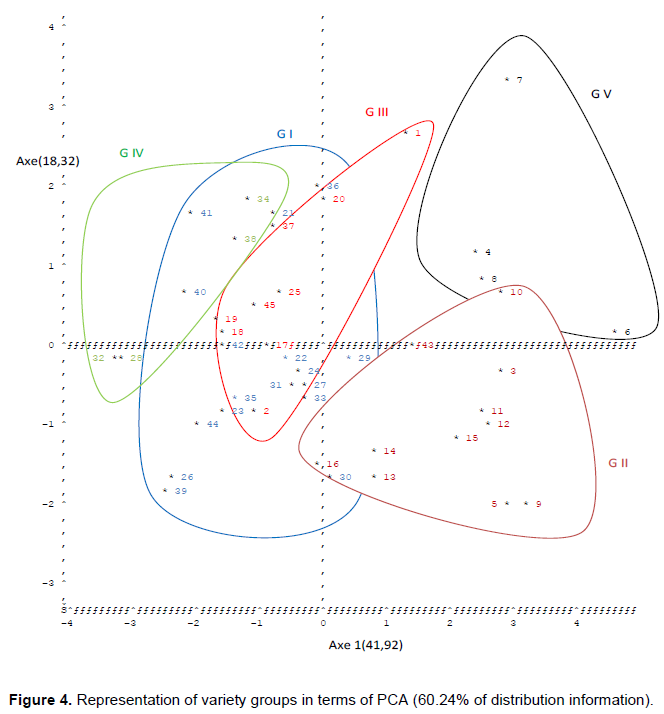
Indeed, all the varieties found on the positive side of the axis1, have the lowest value of the gall index (between 0 and 3) (Appendix 1) and the nematode multiplication rate (PF / PI <1). These varieties, which proved to be resistant five weeks after inoculation, are varieties 4, 8 and 6. Seven of the GV group, varieties 10, 43, 3, 11, 12, 15, 5, 9, 14, 13, 16 of group GII and varieties 29, 30 belong to group GI.
Similarly, all the varieties that are located on the negative side of the axis1 in the plane of projection of the groups given by the ACP (Figure 4), are those which had recorded a fairly high gall index (between 5 and 7); with a multiplication rate of nematodes higher than 1 (PF / PI > 1). These varieties, which had been sensitive after 5 weeks of inoculation, also gave the poor performance of growth and production parameters; they are usually the latest with a high flowering time (more than 50 days after sowing). The susceptible varieties are 41, 21, 40, 42, 22, 31, 35, 23, 44, 26, 41, 24, 27, 39, 25, 33 of the GI group; whereas varieties 37, 25, 45, 19, 18, 17, 2, 1.20 of group GIII and varieties 34, 38, 32, 28 belong to the GIV group.
On axis 2, it is noted that the varieties of group GI, GIII, and GIV previously mentioned in the above paragraph would also be the most sensitive to nematodes because of having recorded higher values of gall index and multiplication rate nematode population; however, this sensitivity is much greater for the varieties found on the positive side of axis 2. These varieties are 34 and 38 of the GIV group; the 41, 40, 21, and 36 varieties of the GI group; and varieties 37, 25, 45, 19, 18 belonging to group GIII.
These susceptible varieties are those that have shown characteristics of sick individuals with low yield with leaf losses, leaf yellowing, and stunting, along with many galls in the roots (Photos 3, 4, 5 and 6).

DISCUSSION
Germination
The seeds of okra are characterized by dormancy of integumentary origin due mainly to slow penetration of water through the seed coat; however, to get a quick lift, it is important to soak the seeds in the water 24 h before sowing. The germination test in a petri dish is very important to see the germinative capacity of the seeds after conservation for a certain time. Regarding the emergence rate, we found that A. esculentus varieties are much later than those of A. esculentus. This is due to the permeability of the seed coat, which is much slower in the varieties of A. caillei, which justifies their late characteristics at the time of germination. Our results are comparable to those obtained by Siemonsma (1979) in the work he had to carry out in Ivory Coast on the cultivation of okra. He had noticed that the guinea type (which is a variety of A. caillei) is generally later than the Sudanese type (A. esculentus variety).
At the exit of the soil, the seedling of the okra is yellowish, then it becomes green immediately after exposure to the light. This green coloration of the cotyledons ensures the first chlorophyllian function before the appearance of real leaves. The cotyledons, therefore, represent the first assimilating leaves. Germination is of the epigeal type.
Ecotypes variability
The study of okra accessions of various origins for their sensitivities relative to the attack of nematodes genus Meloidogyne, showed a strong genetic variability within these varieties. However, it should be noted that even within resistant or susceptible varieties, the variability is rather high not only because the degree of sensitivity differs from one variety to another within the same group defined by the CAH, but also from one group to another.
We can, therefore, accept the hypothesis of inter and intra- varietal variability for the varieties studied. With regard to the varieties which proved to be resistant following the infestation, we can say that these varieties had to carry out a physiological mechanism which remains to be determined, and which had contributed to the reduction of the populations of the inoculated phytoparasites nematodes.
This type of mechanism that some plants do to resist the attacks of plant pests was demonstrated by the works of Singh and Choudhury cited by Huang (1985) in which they show that the varieties of tomatoes attacked by nematodes react to these phytoparasites by slowing down or suppressing the development of these nematodes by the production of phenolic compounds present in large quantities in the tissues of resistant varieties and immune varieties of tomato.
However, it was also noted that among the resistant varieties, only variety 7 of group V had the highest values of the gall index (I Galle = 7) and the multiplication rate (PF / PI = 1.44) nematodes, although it did not show any symptoms of the disease. This assumes that the juveniles counted at its level have very low pathogenicity which is one of the characteristics of the male individuals of these nematodes; in other words, the variety had to secrete a substance which acted on the orientation of the sex of the obtained juveniles and later became male individuals with low pathogenic power. Sarr (1988), Singh and Singh (1988) and Sarr and Prot (1986) noted that juveniles in some resistant plants develop into low pathogenic adult males. So, it is the justification for the reaction of Variety 7 that we evaluated.
This result shows that it is possible to develop improved cultivars and hybrids using appropriate hybridization methods. However, this method is ignored by farmers and is not easily accessible to them. It is therefore important to make farmers aware of the importance of variability within varieties.
Over the last 30 years in Niger, with the successive cycles of drought and the increasing pressure on natural resources, we are witnessing the degradation of ecosystems, the impoverishment of the soil, and also and above all, the disappearance of the number of cultivars leading to genetic erosion. Since then, approaches and strategies have been implemented to ensure their conservation ex and in situ.
Ex situ conservation is carried out in genebanks (e.g. in ICRISAT of Niger where more than 256 varieties of okra are conserved). In situ conservation of agricultural biodiversity is a new technique of conservation in a pleasant environment that values ​​the knowledge of farmers, makes them responsible and understands the scientific basis for the selection and conservation of varieties cultivated and maintained for centuries (IRD, 2004).
West Africa, a zone of origin and domestication of okra, contains a wide genetic diversity that can play a vital role in the development of productive cultivars to meet the growing needs of the populations. Thus, variability within regional ecotypes could be exploited in the development of gene pools with a broad genetic base. For example, we could seek to improve the performance of Niger's accessions so that they can withstand phytoparasites (nematodes for example), by crossing with other accessions of a West African country, presenting stronger resistance characters.
In Niger, the recent research activities of the AVRDC program based at ICRISAT, on the exploitation of hybrid vigor (heterosis), made it possible to select okra varieties adapted to the two seasons (rainy season and dry season). Also, studies on the analysis of the morphological evolution and genetic diversity of okra varieties conducted by Siemonsma and Hamon (2004), and those conducted by Mahamadou et al. (2005) on water stress revealed changes. Morphological differences between the varieties studied showed significant variation in terms of diversity.
The genetic material that has just been characterized in this study will be used in the heterotic grouping of West African okra. Thus, by crossing individuals from these groups that are quite divergent, we intend to develop a better performance because of the high rate of heterozygosity and therefore the hybrid vigor (exploitation of heterosis).
Relationship between the measured characters
The correlations observed between the measured parameters and the principal components give proof of the existence of associations between certain characters.
For the most significant, we note that the most resistant varieties have a gall index and nematode multiplication rate quite low or zero and are generally the earliest, which ensures a good development of other growth parameters (height of the plant) and productions (fresh weight and aerial dry weight). These varieties are grouped into the GI, GV and GII groups based on the hierarchical ascending classification (CAH).
Varieties grouping
Our varieties are divided into five groups. The gall index and the nematode multiplication rate are the most discriminating traits for the groups of varieties formed by C.A.H. This enabled to characterize the different groups that proved to be resistant or sensitive five weeks after inoculation in the following.
The varieties that make up groups II, V and only two varieties in group I, namely 29 and 30, are all nematode-resistant because they are associated with the positive side of axis 1, which has low values ​​of the index, gall and multiplication rate of nematodes. These varieties are therefore characterized by low values ​​of the gall index (ranging from 0 to 3) and the nematode multiplication rate (PF / PI >1). They also have fresh weight efficiencies and much higher aerial dry weight with important values ​​for height. These characteristics of the resistant varieties of okra have been demonstrated in the work done by Khan et al. (1997), in which reduction of the multiplication rate of nematodes of the species M. javanica on the roots of resistant varieties of okra is shown. The resistant varieties of these groups are also early. When we look at the species to which these varieties belong, we find that they are the varieties of the species A. esculentus, except the variety 43 of group II which belongs to the A. caillei.
The varieties that make up the other groups, that is, groups III, IV and I with the exception of varieties 29 and 30, were found to be susceptible to nematodes of the genus Meloidogyne, as they were associated with the negative side of axis 1 which has high values ​​of gall index and nematode multiplication rate. These varieties are therefore characterized by high values ​​of gall index and nematode multiplication rate (PF / PI >1). They also have low yields in fresh weight and aerial dry weight with low value for height. These susceptible varieties at five weeks after inoculation are also late. Our results are comparable to the results obtained by Das and Sinha (2005), in which they showed that 27 cultivated varieties of okra were susceptible to the attack of M. incognita nematodes. The same is true for the work done by Naidu et al. (2006), in which three sensitive varieties of okra were determined after examining a number of plant varieties for the M. incognita attack. Among these determined varieties, they announced that the variety cv. Arka Anamica was highly sensitive with a gall index and a fairly high nematode multiplication rate (PF / PI >1), whereas the variety cv. Prabhani Kranti (PI548700) was moderately tolerant.
Group characterization
Starting from the results obtained on the characterizations of the different groups of varieties we can say that the discrimination of these groups is made essentially on the basis of the gall index (I Galle) and the nematode multiplication rate (PF / PI). Results of the analysis of variance performed on the measured parameter means show that the groups differ only in height, aerial dry weight (Psaer), aerial weight (Pfaer), flowering time (FLO) and to a lesser extent the fresh weight and root dry weight (Pfrac) and (Psrac).
However, it has been noted that Group I comprise only two resistant varieties (29 and 30), all belonging to A. esculentus. These are the earliest varieties of the group. The other varieties of group I are late and sensitive. Group II contains varieties that are all resistant. These varieties, which are 10, 3, 11, 12, 15, 5, 9, 14, 13, and 16 are all early. Group III consists of susceptible and late varieties with the exception of variety 1, which is early. It was also found that variety 1, although sensitive to nematodes, also had a high yield in fresh weight and dry root weight. This assumes that galls developed at its level are much larger than those developed for other susceptible varieties. Group IV comprises only late and sensitive varieties because they had the highest values ​​of gall index and nematode multiplication rate as shown by their projection in the plane defined by the PCA. Group V consists of varieties that are all resistant and early. These varieties gave good yields at the measured parameters except for the gall index and the rate of multiplication of the nematodes which are very low and characterizing the resistance capacity in these varieties.
CONCLUSION
The present study has allowed us to highlight the existence of the great diversity of okra genetic resources in West Africa and also in other continents of the world. The variability observed within the varieties indicates that there are at least groups which are different from each other on the basis of quantitative and qualitative characteristics obtained five weeks after inoculation.
Screening of 48 varieties of okra of various origins showed that resistant varieties show significant genetic variability for nematode resistance. The resistant varieties of the trial showed a reduced number of nematode multiplication rates with low gall index values ​​at their roots, and also relatively higher yields for the other measured parameters. They represent important genetic sources for the resistance of okra to nematodes. For better accuracy of parasite resistance, it is desirable to identify alleles for resistance and to fix them. In the end, we believe that this study can serve as an important contribution for better valorization of the okra genetic resources in the breeding programs.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Alake CO, Ariyo OJ (2012). Comparative Analysis of Genotype x Environment Interaction Techniques in West African Okra, (Abelmoschus caillei, A. Chev Stevels). Journal of Agricultural Science 4(4):135-150. |
|
|
Agbo AE, Gnakri D, Beugré GM, Fondio L, Kouamé C (2008). Maturity degree of four Okra fruits varieties and their nutrients composition. Electronic Journal of Food and Plants Chemistry 3(1):1-4. |
|
|
Dabiré-Binso CL, Ba MN, Somé K, Sanon A (2009). Preliminary studies on incidence of insect pest on okra, Abelmoschus esculentus (L.) Moench in central Burkina Faso. African Journal of Agricultural Research 4(4):1488-1492. |
|
|
Das N, Sinha AK (2005). Reaction of okra varieties to root-knot nematode (Meloidogyne incognita). Indian Journal of Nematology 35:214. |
|
|
DS/MDA (2008). Annuaire des statistiques, Ministère du Développement Agricole. Niamey, Niger 128 p. |
|
|
Food and Agriculture Organization (FAO) (2004). Handbook on Human Nutritional Requirements. FAO, Rome, Italy. |
|
|
Fondio L, Kouame C, Djidji AH, Traore D (2003). Effet de la date de semis sur la production du gombo (Abelmoschus spp.) dans le centre de la cote d'ivoire. Agronomie Africaine 15(1):13-27. |
|
|
Fondio L, Kouame C, Djidji AH, Traore D (2011). Caractérisation des systèmes de culture intégrant le gombo dans le maraîchage urbain et périurbain de Bouaké dans le Centre de la Côte d'Ivoire. International Journal of Biological and Chemical Science 5(3):1178-1189. |
|
|
Glegle KR, Kokode GG (2004). Techniques statistiques univariées et multivariées : applications sur ordinateur. Institut National des recherches Agricoles du Bénin /DBAM/PISB/CRA-Agonkanmey/ INRAB 68 p. |
|
|
Hamon S (1988). Organisation Evolutive du Genre Abelmoschus (Gombo): Co-Adaptation et Evolution de Deux Espèces de Gombo Cultivées en Afrique de l'Ouest (Abelmoschus esculentus et Abelmoschus caillei). Institut Français de Recherche Scientifique pour le Développement en Coopération 46 :19-21 |
|
|
Haougui A, Kollo AI (2008). Les nématodes ennemis invisibles des maraichers. |
|
|
Haougui A (1999). Les Nématodes Parasites des Cultures Maraichères au Niger: Importance et Méthodes de Lutte par Utilisation de Plantes. Thèse de Doctorat du CARFOP/ Université de Dschang Cameroun 114 p. |
|
|
Huang JS (1985). Mechanisms of résistance to root-knot nématodes. Dans ¨An Advanced Treatise on Meloidogyne. Biology and Control¨ SASSER, J.N., CARTER, C.C., eds., Raleigh, North Carolina State University Graphics, USA 1:165-174. |
|
|
Idi-Saidou S, Moustapha G, Kumar S, Bakasso Y, Ali M (2019). Regeneration and Preliminary Characterization of Asian and African Okra (Abelmoschus spp.) Germplasm. European Scientific Journal 15(6):21-32. |
|
|
INERA/BF (2001). Rapport Technique Annuel. Coordination Nationale du Projet In Situ. INERA, Ouagadougou, Burkina Faso 110 p. |
|
|
IRD (2004). Rapport Annuel d'Activité de l'IRD, Niamey, Niger 20 p. |
|
|
Khan MW, Khan AA, Khan MR (1997). Effect of soil salinity on penetration, development and pathogenicity of Meloidogyne incognita on okra and cucumber. Indian Journal of Nematology 27:194-208. |
|
|
Kumar S, Chowdhury S (2019). Okra Breeding: Recent Approaches and Constraints. Annals of Biology 35(1):55-60. |
|
|
Mahamadou S, Gerard Z, Didier B (2005). Expression de différents écotypes de gombo (Abelmoschus esculentus L.) au déficit hydrique intervenant pendant la boutonnisation et la floraison. Biotechnology, Agronomy and Society and Environment 10(1):43-54. |
|
|
Naidu PH, Haritha V, Sudheer MJ (2006). Screening of vegetable crop varieties for resistance to root-knot nematode (Meloidogyne incognita). Indian Journal of Nematology 36:293-229. |
|
|
Ouédraogo ZA (2009). Caracterisation agromorphologique comparé de cinq varietés de gombo (Abelmoschus esculentus L. Moencb). Mémoire présenté à l'université polytechnique de Bobo-Dioulasso, Burkina Faso, pour l'obtention du diplôme d'Ingeniorat 61 p. |
|
|
Ouédraogo MH (2016). Etude de la diversité génétique des gombos [Abelmoschus esculentus (L.) Moench] cultivés au Burkina Faso. Thèse Doctorat, Université Ouaga I Pr KI-ZERBO 149 p. |
|
|
Ratnadass A, Ousmane ZM, Hame AKK, Sanjeet K, Isabelle G, Philippe R, Halarou S, Moussa A, Abdul AM, Saïdou AS, Issoufou S (2014). Potential of pigeon pea as a trap crop for control of fruit worm infestation and damage to okra. Agricultural and Forest Entomology pp. 1- 8. |
|
|
Sani IS (2010). Criblage de genotypes de gombo (Abelmoschus spp.) Pour leur resistance aux nematodes (Meloidogyne spp.). Diplôme d'Etudes Approfondies (D.E.A) en Biologie Appliquée, Option : « Protections et Restaurations des Ecosystèmes Soudaniens et Sahélo-Sahariens » Université Abdou Moumouni de Niamey 77 p. |
|
|
Sanwal SK, Singh M, Singh B, Naik PS (2014). Meeting report: Resistance to yellow vein mosaic virus and okra enation leaf curl virus: Challenges and future strategies. Current Science 106:1470-1471. |
|
|
Sanwal SK, Venkataravanappa V, Singh B (2016). Resistance to bhendi yellow vein mosaic disease: A review. Indian Journal of Agricultural Sciences 86:835-843. |
|
|
Sarr E (1988). Nématologie Tome 11. Département de Formation en Protection des Végétaux. Centre AGRHYMET, Niamey, Niger 82 p. |
|
|
Sarr E, Prot JC (1986). Pénétration et développement des juvéniles d'une souche de Meloidogyne javanica et d'une race B de M et d'une race B de M. incognita dans les racines du fonio (Digitaria exilis Stapf). Revue Nématology 8:59-65. |
|
|
Seinhorst JW (1950). De betekenis van de toestand von de grand voor het optreden van. Aanstasting door het stengelaaltje (Ditylenchus dispsaci (Kühn) Filipjev). Tijdrchr. Plziekt 56:292-349. |
|
|
Siemonsma JS (1979). La Culture du Gombo (Abelmoschus) Légume-Fruit Tropical avec Référence Spécial a la Cote d'Ivoire, 120p. |
|
|
Siemonsma JS, Hamon S (2004). Abelmoschus esculentus L. Moencb. In : Ressources vegetales de l'Afrique Tropicale 2. Fondation PROTZ. Wageningen Pays-Bas pp. 25-30. |
|
|
Singh HP, Singh V (1988). Evaluation of oilcakes and nématicides of Meloidogyne incognita infecting eggplants. Indian Journal of Nematology 18:366-368. |
|
|
Zeck WM (1971). A rating scheme for field evaluation of root-knot nematode infestations. Aanstasting door het stengelaaltje (Ditylenchus dispsaci (Kühn) Filipjev). Tijdrchr. Plziekt 56 :292-349. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


