Short Communication

ABSTRACT
INTRODUCTION
Legumes can form a nitrogen fixing symbiosis with soil bacteria called rhizobia (the RL symbiosis). They can also like most plants, form symbiotic associations with arbuscular mycorrhizal (AM) fungi, which facilitate plants’ phosphate nutrition. In both interactions, the symbionts are hosted inside the plant root. Nitrogen-fixing rhizobia are housed in intracellular symbiotic structures within nodules, while AM fungi form intracellular symbiotic structures called arbuscules, within cortical root cells and can also develop external mycelium which extends from around the root. Molecular genetics studies performed on the model legumes Medicago truncatula and Lotus japonicas have shown that the establishment of the nodulation and the mycorrhization processes share a common signaling pathway, required for the initiation of endosymbiotic programs in host plants. In M. truncatula, three genes called the DMI genes (which does not make infection) are involved in this pathway (Catoira et al., 2000). The common signaling pathway diverges after DMI3 which represents the last known gene common to both symbiosis. DMI3 encodes calcium and calmodulin depen-dent protein kinase (CCaMK), located in the nucleus, and is supposed to perceive and transduce calcium signals generated upon perception of the symbiotic signals.
Therefore, it has been hypothesized that DMI3, depending on its activation determined by the calcium signature, could phosphorylate substrates involved either in nodula-tion or mycorrhization (Levy et al., 2004).CCaMKs has been studied in other plants and its activity depends on an auto-inhibitory domain that negatively regulates the kinase activity. Removal of this domain or point mutations at the auto-phosphorylation site respect-tively, in M. truncatula and L. japonicus, leads to a con-stitutively-active form of CCaMK that can induce spon-taneous symbiotic responses and nodulation in the absence of Nod factors (Gleason et al., 2006; Tirichine et al., 2006). Otherwise, a protein interacting with DMI3, named IPD3 (interacting protein of DMI3) in M. truncatula and CYCLOPS in L. japonicus, has been identified (Messinese et al., 2007; Yano et al., 2008). CYCLOPS, which is phosphorylated in vitro by the CCaMK seems to be important for the infection process in both symbioses, but is dispensable for nodule organogenesis, suggesting that CCaMK has different molecular targets during the nodulation process. Recently, Rival et al. (2012, 2013) showed that both rhizobial and mycorrhizal symbiosis are controlled by DMI3 in a cell autonomous way. Nodule organogenesis was not observed when DMI3 expression was restricted either to the epidermis or to the cortex, but was restored when DMI3 was expressed in both tissues. Moreover, it has been shown that transcription factor NIN downstream DMI3, negatively regulates infection but positively regulates nodule organogenesis during the course of the symbiosis (Yoro et al., 2014). Through all these observations, the infection process needs the full activation of CCaMK via the calcium spiking generated by the common signalling pathway and possibly an additional calcium signal. In this context, the use of a constitutively-active form of DMI3 could be a valuable tool to search for downstream targets which could potentially include further components of the common symbiotic signaling pathway or components specific to Nod or Myc signalling. From previous work, DMI3 variant (DMI3-311) in terms of production was described as a tool for identifying substrates potentially involved in nodulation or mycorrhization (Kassouar and Baba Hamed, 2011). In this study, we verified the functionality of this variant and its ability to induce spontaneous nodules in the M. truncatula dmi3 mutant.
MATERIALS AND METHODS
Enzymatic radio activity assay
The catalytic activity of the recombinant DMI3-311 was examined by a test of auto-phosphorylation of the protein. Indeed, a volume of 40 μL purified fraction was added to a reaction buffer (10 mM Mg Cl2, 50 µM [γ-33P] ATP, 1mM DTT and 10 mM MnCl2), the mixture was then incubated at 25°C for 1 h. After centrifugation, the labeled proteins were separated by SDS-PAGE gel electrophoresis (10% acrylamide). The labeling of phosphorylated proteins was revealed on X-Ray film (Amersham) after 4 to 5 days of exposure at -80°C.
M. truncatula transformation by Agrobacterium rhizogenes
Transformation of dmi3 mutant by Agrobacterium rhizogenes using pCambia2202 binary vector with DMI3 1-311 construction (the tagged form: StrepTagII and no tagged form were used separately) under control of its own promoter (pDMI3-DMI3 1-311) (Godfroy et al., 2008), began with (TRV25 allele) dmi3 mutant germination. After the step of scarifying by H2SO4 (95%), surface-sterilised seeds were sown on agar plates and placed for 3 days in the dark at 4°C then left overnight at 25°C to germinate. After approximately 30 h germination, when seedlings had a radicle length of approximately 10 mm, the radicle was sectioned approximately 3 mm from the root tip with a sterile scalpel. Sectioned radicles were inoculated by coating the freshly cut surface with A. rhizogenes grown on TY solid medium (Boisson-Dernier et al., 2001). Thereafter, the inoculated sectioned seedlings were placed on slanted agar (Laboratoire Industriel de Biologie, Avignon, France) containing a modified Fahraeus medium supplemented with kanamycin (1mM CaCl2, 0.5 mM MgSO4, 0.7 mM KH2PO4, 0.8 mM Na2HPO4, 50 μM FeEDTA, 0.5 mM NH4NO3, supplemented with 0.1 mg of MnSO4, CuSO4, ZnSO4, H3BO3, and Na2MoO4 per liter) in square Petri dishes (12 × 12 cm). After several incisions in the Parafilm seal allowing gas exchange, the Petri dishes were placed vertically in a growth chamber at 20°C for 1 week (16-h photoperiod and a light intensity of 70 μE/s/m2) to optimize the transformation frequency, and then transferred to a 25°C growth chamber (identical light conditions) for 2 weeks.
For nodulation assays, transformed plants were transferred separately in two different environments: nto growth pouches, with 7 ml of Fahraeus liquid medium (with nitrates) (3 to 4 plants/pouch), and incubated at 25°C for 3 to 4 weeks; and to sepiolite (Agrauxine, Quimper)/sand (2:1 volume mix) pots and grown at 25°C with 18 h light/6 hour dark cycles (Catoira et al., 2000).
Microscopy methods
The presence of spontaneous nodules was first observed by optical microscope (immersion x100) (Zeiss Axiophot, Carl Zeiss, Germany). Transformed roots were cleared with 3% sodium hypochlorite for 15 min and observed with a Leica MZFLIII stereomicroscope (Leica Microsystems, Wetzlar, Germany).
RESULTS AND DISCUSSION
In order to identify targets DMI3, a biochemical study based on obtaining a truncated form of DMI3 displaying constitutive activity was chosen to overcome stimulation by Nod factors and providing DMI3 homogeneous samples that would be permanently in its interaction with potential targets. For this we followed Gleason et al. (2006) who showed that expression of a truncated form of DMI3 in the roots of M. truncatula induces symbiotic responses in the absence of symbiont. These researchers present the construction of a set of partially-deleted mutants to DMI3, 1-326 and 1-311, corresponding to the kinase domain without two regulatory domains (visinin domain and central domain). In vitro phosphorylation tests on artificial substrate showed constitutive activity, independent calcium, 1-326 and 1-311 forms of DMI3, corresponding to about 30 and 40%, respectively of the activity of the whole protein in the presence of calcium and calmodulin. From previous work done by authors, a truncated form of DMI3 was produced, the variant DMI3-311. Here, the catalytic activity of this variant was first verified, in vitro, by autophosphorylation assay of the protein.
Catalytic activity of the DMI3-311 protein
The presence of auto-phosphorylation was detected using a radioenzymatic labeling of the protein with [g-33P] ATP. It is important to know that at the end of the N-terminal kinase domain is found the binding domain to ATP and at its C-terminal the Thr267, auto-phosphorylation site of the protein (Sathyanarayanan et al., 2000; Sathyanarayanan and Poovaiah, 2002). Figure 1 reveals the presence of phosphorylation at the expected molecular weight about 39 to 42 kDa confirming that the DMI3-311 recombinant protein is active.
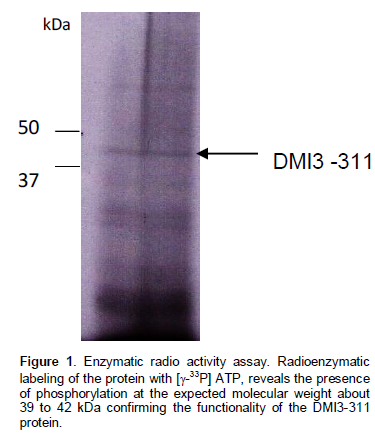
Spontaneous nodulation
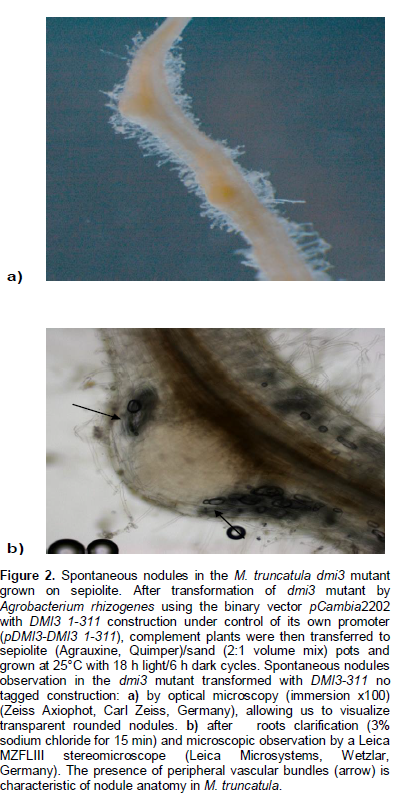
We tested, in planta, the ability of the variant DMI3-311 to induce spontaneous nodules in absence of bacterial symbiont and Nod factor. For this, we performed tests of complementation of dmi3 mutant using the binary vector pcambia2202 with DMI3 1-311 construction under control of its own promoter (pDMI3-DMI3 1-311) (Godfroy et al., 2008) (the tagged form: Strep TagII, and no tagged form were used separately) by Agrobacterium rhizogenes transformation. Composite plants were then transferred separately in two different environments, into growth pouches with Fahraeus liquid medium and to sepiolite pots. The transformation efficiency, in absence of any stimulation symbiotic (Nod factors or bacterial symbiont) results first, by the early nodulin ENOD11gene expression conferring a blue coloration to root epidermic tissue (data not shown) and the formation of structures with the anatomy of a nodule. Few spontaneous nodules were observed in plants complemented with the no tagged DMI3 1-311 construction and grown in sepiolite (Figure 2). Microscopic observations revealed transparent rounded nodules (Figure 2a) characterized by the presence of peripheral vascular bundles characteristic of nodule anatomy in M. truncatula (Figure 2b). These results were confirmed after histological assay (data not shown). However, no nodules were observed in plants complemented with the StrepTagII DMI3 1-311 construction.
In light of these results, we can claim to be in possession of a constitutively active truncated form of the kinase DMI3, this molecular tool will allow us to achieve phosphorylation in vitro, first in extracts of cell nuclei suspensions of M. truncatula A17 (wild strain) and the mutant dmi3 (as suspension cells are easy to grow) and in extracts of root, in order to identify substrates or DMI3 molecular partners involved in the process of nodulation or mycorrhiza. Preliminary in vitro phosphorylation studies, on cell suspensions nuclei extracts of M. truncatula A17 (wild type) and dmi3 mutant are encouraging but need to be confirmed.
CONCLUSION AND RECOMMENDATION
By decoding calcium signals, DMI3/CCaMK could play a central role in orientating the signalling pathway leading to nodulation or mycorrhization. The possibility to produce active DMI3-311 gives the opportunity to search for its substrates by performing in vitro phosphorylation assays using plant extracts.
CONFLICT OF INTERESTS
The author(s) have not declared any conflict of interests.
ACKNOWLEDGMENTS
S. Kassouar is grateful for the help rendered by Drs J. J. Bono, C. Rosenberg and G. Borderies during her stay in France in the frame of her research which were supported by the Algerian Ministry of Higher Education and Scientific Research.
REFERENCES
|
Boisson-Dernier A, Chabaud, M, Garcia F, Bécard G, Rosenberg C, Barker DG (2001). Agrobacterium rhizogenes-transformed roots of Medicago truncatula for the study of nitrogen-fixing and endomycorrhizal symbiotic associations. Mol. Plant Microbe Interact.14 (6):695-700. Crossref |
||||
|
Catoira R, Galera C, de Billy F, Penmetsa RV, Journet EP, Maillet F, Rosenberg C, Cook D, Gough C, Dénarié J (2000). Four genes of Medicago truncatula controlling components of a Nod factor transduction pathway. Plant Cell 12:1647-1666. Crossref |
||||
|
Gleason C, Chaudhuri S, Yang T, Munoz A, Poovaiah BW, Oldroyd GED (2006). Nodulation independent of rhizobia induced by a calcium-activated kinase lacking autoinhibition. Nature 441:1149-1152. Crossref |
||||
|
Godfroy O, Debelle F, Timmers T, Rosenberg C (2006). A rice calcium- and calmodulin-dependent protein kinase restores nodulation to a legume mutant. Mol. Plant Microbe Interact.19:495-501. Crossref |
||||
| Kassouar S, Baba Hamed MB (2011). Heterologous expression and partial purification of calcium and calmodulin dependant protein kinase (CCaMK) of Medicago truncatula. Afr. J. Biotechnol. 10(61):13189-13193. | ||||
|
Levy J, Bres C, Geurts R, Chalhoub B, Kulikova O, Duc G, Journet EP, Ane JM, Lauber E, Bisseling T, Dénarié J, Rosenberg C, Debellé F (2004). A putative Ca2+ and calmodulin-dependent protein kinase required for bacterial and fungal symbioses. Science 303:1361-1364. Crossref |
||||
|
Messinese E, Mun JH, Yeun LH, Jayaraman D, Rouge P, Barre A, Lougnon G, Schornack S, Bono JJ, Cook DR, Ane JM (2007). A novel nuclear protein interacts with the symbiotic DMI3 calcium and calmodulin-dependent protein kinase of Medicago truncatula. Mol. Plant Microbe Interact. 20:912-921. Crossref |
||||
| Rival P, Bono JJ, Gough C, Bensmihen S, Rosenberg C (2013). Cell autonomous and non-cell autonomous control of rhizobial and mycorrhizal infection in Medicago truncatula. Plant Signal. Behav. 8(2): e22999-1- e22999-4. | ||||
|
Rival P, De Billy F, Bono JJ, Gough C, Rosenberg C and Bensmihen S (2012). Epidermal and cortical roles of NFP and DMI3 in coordinating early steps of nodulation in Medicago truncatula. Development 139:3383-339. Crossref |
||||
|
Sathyanarayanan PV, Cremo CR, Poovaiah BW (2000). Plant chimeric Ca2+/calmodulin-dependent protein kinase. J. Biol. Chem. 275: 30417-30422. Crossref |
||||
|
Sathyanarayanan PV, Poovaiah BW (2002). Autophosphorylation dependent inactivation of plant chimeric calcium/calmodulin-dependent protein kinase. Eur. J. Biochem. 269:2457-2463. Crossref |
||||
|
Tirichine L, Imaizumi-Anraku H, Yoshida S, Murakami Y, Madsen L H, Miwa H, Nakagawa T, Sandal N, Albrektsen A, Kawaguchi M, Downie A, Sato S, Tabata S, Kouchi H, Parniske M, Kawasaki S, Stougaard J (2006). Deregulation of a Ca2+/calmodulin-dependent kinase leads to spontaneous nodule development. Nature 441:1153-1156. Crossref |
||||
|
Yano K, Yoshida S, Müller J, Singh S, Banba M, Vickers K, Markmann K, White C, Schuller B, Sato S, Asamizu E, Tabata S, Murooka Y, Perry J, Wang T L, Kawaguchi M, Imaizumi-Anraku H, Hayashi M, Parniske M (2008). CYCLOPS, a mediator of symbiotic intracellular accommodation. Proc. Natl. Acad. Sci. 105:20540-20545. Crossref |
||||
|
Yoro E, Takuya S, Koichi T, Hikota M, Hidehiro F and Masayoshi K (2014). A Positive Regulator of Nodule Organogenesis, NODULE INCEPTION, Acts as a Negative Regulator of Rhizobial Infection in Lotus japonicas. Plant Physiol. 165:747-758. Crossref |
||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


