Full Length Research Paper

ABSTRACT
INTRODUCTION
The conservation of crops in the world crucially depends on the knowledge of their genetic diversity. Molecular markers, such as simple sequence repeats (SSRs), reveal diversity at the DNA level; and thus provide a fundamental tool for germplasm conservation and genotyping. Avocado is one of the highly marginised crops in Ghana though it has a very high nutritive value and a good potential for commercial production (Campbell and Malo, 1976; Morton, 1987; Verheij and Coronel, 1991; FAOSTAT Database, 2001). There have been very few studies that investigated alternative ways of improving the production and conservation of Avocado in Ghana, in contrast to the increasing attention given to the crop elsewhere (Knight, 2002). Consequently, we are witnessing indiscriminate cutting of this important resource in Ghana due to the low profile given to it.
Traditionally, morphological characters are used to identify the crop for selection and breeding. However, the efficiency of morphological traits has been mixed. Whilst morphological traits have proved useful in distinguishing between species in many cases (Rhodes et al., 1971; Morton, 1987), recent studies reveal that morphological characterisation is not always very informative because the traits can be altered by environmental conditions and thus mislead species identification (Bergh et al., 1973; Furnier et al., 1990; Gutiérrez-Diez et al., 2009). The use of model genetic systems in plant and animal studies nowadays represent a more reliable alternative way that greatly increases our understanding of how genomes regulate phenotype.
The avocado plant, Persea americana Mill. (Lauraceae), is indigenous to Central and South America notably Mexico and has developed varieties, which have adapted to a wide range of climatic conditions including those of Asia and Africa (Bergh, 1969; Rhodes et al., 1971). Three botanical varieties of avocado have traditionally been recognised; namely, Mexican, Guatemalan and West Indian. These varieties are distinguishable on the basis of morphological, physiological, and horticultural traits; and they are adapted to different climates and ecological conditions (Bergh, 1995; Bergh and Lahav, 1996). They also correspond to the varieties P. americana var. drymifolia (Schlecht. et Cham.) Blake, P. americana var. guatemalensis Williams. and P. americana var. americana Mill., respectively, (Bergh et al., 1973; Bergh, 1995). Several attempts have been made to refine the classification of avocado such as numerical taxonomical methods based on morphological characters (Rhodes et al., 1971). Such methods revealed that cultivars tended to cluster into three groups, representing three varieties (Rhodes et al., 1971).
Further studies applied genetic-based approach such as restriction fragment length polymorphism (RFLP) of chloroplast DNA, ribosomal DNA, and the genes coding for the enzyme cellulase to infer a phylogeny of avocado (Furnier et al., 1990). In that instance, the results lent some support to the current classification which placed the varieties drymifolia, guatemalensis and americana in a single species P. americana (Bergh et al., 1973). More recent studies have used the markers of minisatellite and microsatellite DNA to characterise and differentiate botanical varieties of avocado (Lavi et al., 1991; Mhameed et al., 1997; Acheampong et al., 2008).
Microsatellites are said to be highly polymorphic and useful as genetic markers that have been used in defining genetic similarities in crops such as maize, sorghum and wheat (Röder et al., 1995; Smith et al., 1997; Taramino et al., 1997; Uptmoor et al., 2003; Menz et al., 2004). While both DNA fingerprints (DFP) and SSR markers were used to estimate the heterozygosity level in the avocado genome and to define genetic relationships in the Persea genus (Mhameed et al., 1996; 1997), none of these markers has so far been used to construct a genetic linkage map of the avocado genome (Sharon et al., 1997) and this does not give a pictorial view of the relationships within avocados. Moreover, most of the research done on avocados are concentrated in the developed world like the United States of America and Israel (Morton, 1987; Furnier et al., 1990; Schnell et al., 2003), whilst we know very little about their genetic diversity in the developing and third world countries like Ghana. Our study aimed at defining the genetic variations between avocado accessions in Ghana using microsatellite. We therefore used the potential of SSRs to track pollen movement (Queller et al., 1993; Jarne and Lagoda, 1996; Goldstein and Pollock, 1997; Sunnucks, 2000) and their suitability for studying genealogical relationships (Goldstein et al., 1995) to characterise the avocados in Ashanti and Central Regions of Ghana for the first time.
MATERIALS AND METHODS
We sampled materials from healthy leaves of 71 avocado plants in the Ashanti and Central Regions of Ghana, as well in the vicinity of the Cocoa Research Institute of Ghana (CRIG) in New Tafo in the Eastern region of Ghana. Thirteen avocado plants out of the total of 71 used were from the avocado farm of CRIG. The 13 plants from the CRIG avocado farm were mostly of West Indian (P. americana var. americana.) and the Guatemalan (P. americana var. guatemalensis Williams) varieties. These were used as controls in this study.
Genomic DNA extraction
DNA was extracted following a modified cetyltrimethyl ammonium bromide (CTAB) protocol (Aldrich and Cullis, 1993). Total DNA was washed in 1 ml washing buffer made of 76% ethanol and 10 mM ammonium acetate, and again in 1 ml ethanol (80%). It was then stored at -20?C. Two microliters of each of the genomic DNA samples was dissolved in 3 µl of TE [1 mM Tris HCl (pH 8), 0.1 m M EDTA (pH 8)] buffer and 1 µl of 6X sample buffer (Bromophenol blue and xyelne cyanol) was added. We used 25 ng/µl of DNA for polymerase chain reaction (PCR).
SSR markers
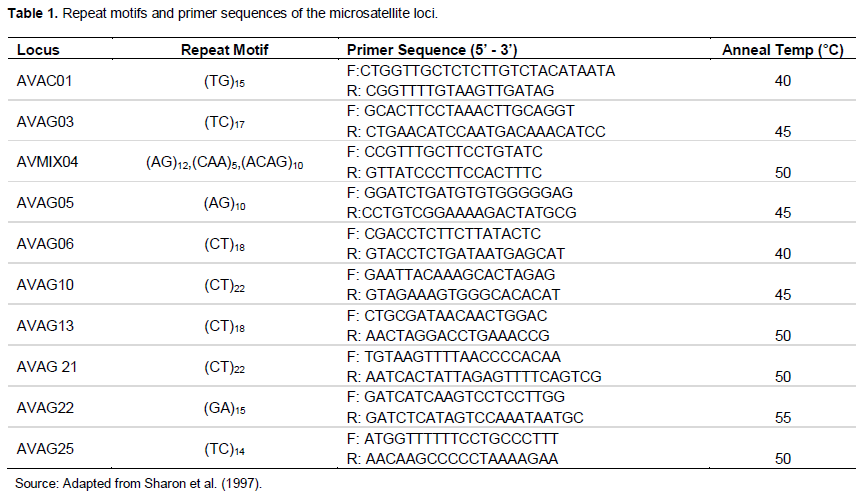
Ten (10) pairs of microsatellite primers designed by Sharon et al. (1997), and later applied by Schnell et al. (2003) (Table 1) were used for the PCR.
PCR amplification and electrophoresis
We performed amplifications using 16 to 50 ng of genomic DNA as template, 0.5 µM of each of the forward and reverse primers, in a 10 µl reaction volume using the AccuPowerTM PCR PreMix (USA Bioneer Inc., Alameda, USA) (DNA polymerase, DNTPs, a tracking dye, and reaction buffer in a premixed format). The PCR reaction was carried out on 2720 Thermal Cycler (Applied Biosystems, Singapore) and consisted of the following cycle: 3 min denaturation at 94°C, followed by 40 cycles of denaturation at 94°C for 30 s, 1 min at appropriate primer annealing temperature (Table 1), 1 min extension at 72°C. The amplification finished with an extension at 72°C for 10 min, followed by maintenance of the reaction mixture at 4°C at infinity until removed for storage at -20°C.
Electrophoresis
Electrophoresis was done using 3 µl of denaturing buffer (95% formamide, 0.02 M EDTA pH 8, 1% bromophenol blue, 1% xylene cyanol, 10 mM NaOH) added to 3 µl of the PCR products. Equal volume of the denaturing buffer was added to 3 µl DNA ladder (10 bp ladder, diluted to 0.1 µg/µl in doubled distilled water). The mixture was denatured at 95°C for 5 min and then immediately chilled on ice. Each was loaded into a well of a 49-well plate (4 mm thick) for electrophoresis on a DNA sequencing gel containing 6% polyacrylamide, 8 M urea and 1 X TBE (Tris-Boric acid-EDTA buffer). Gels were run at 100 W or V constant power and 2 kV for 2 to 2.5 h, using a Bio-Rad Sequi-Gen® GT Nucleic Acid Electrophoresis Cell (Bio-Rad, Consult EG 261, Belgium) and power pack (Bio-Rad Power Pac 300) (Bio-Rad, Consult EG 261, Belgium) and 1 X TBE as running buffer. The products were visualised by silver staining using the method described by Bassam et al. (1991).
Data analysis
For each gel, the distance migrated by each marker size of the DNA ladder was measured using a ruler. A line graph of the distance travelled by the marker size was plotted for each gel; the equation of the relationship between them was then used to estimate the size of the unknown SSR bands of the PCR products. Allele size and the total numbers of alleles were determined for each SSR locus. Bands for same SSR locus with different molecular weight were scored as alleles. The scored alleles were coded using FlexiBin (Bill Amos, Cambridge, UK; http://www.zoo.cam.ac.uk/departments/molecularecology/FlexiBin.pdf) analysis. Gene diversity values for each locus and the average across all loci for all populations were calculated using Nei’s unbiased estimate (Nei, 1973). The numbers of alleles and the allelic frequencies for each SSR, and across all populations as well as the unbiased gene diversity (Hnb), and the observed hetero-zygosity (Hobs), for all populations were estimated using PowerMarker version 3.25 (Liu and Muse, 2004). A phylogenetic tree was constructed for all 71 individuals genotyped following the Unweighted Paired Group Method (UPGMA) (Sneath and Sokal, 1973). Following this, dendrograms of genetic relationships among the genotype studied were produced using PHYLIP version 3.5 and TreeView version 1.6.6 (Saitou and Nei, 1987; Felsenstein, 1989).
RESULTS
DNA analysis and microsatellite typing
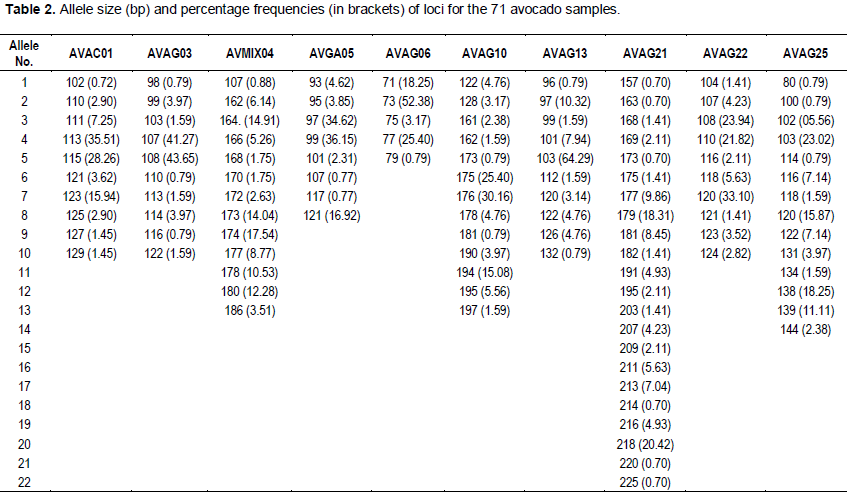
The 10 microsatellites loci had varying degrees of polymorphism, generating 115 alleles across the population sampled (Table 2). The number of alleles varied from five in AVAG06 to 22 in AVAG21; with an average of 11.5 alleles per locus. All SSR loci used were polymorphic. The average gene diversity was 0.7529 (range = 0.5636 to 0.8907) (Tables 3 and 4). The number, size and frequencies of alleles, observed hete-rozygosity (Hobs) number of homozygotes, heterozy-gotes and null alleles across all loci are summarised (Table 3). Allele size ranged from 71 (AVAG06) to 225 bp (AVAG21), with allele frequencies varying between 0.70 (AVOC01) and 64.29% (AVGA13) (Table 2). The allelic frequencies at the different SSR loci varied significantly (Table 3). Across all the accessions screened, 336 (47.3%) loci were homozygous, while 312 (43.9%) loci were heterozygous. The highest Hobs (Hobs = 0.6761) was recorded at the AVAG21 locus, whereas the lowest (Hobs = 0.3333) occurred at the locus AVAG10. The overall mean Hobs of all populations, calculated across all loci was 0.4765. A total of 180 genotypes and 115 alleles were detected; with a mean of 11.5 per locus (Table 4). Unbiased gene diversity (Hnb) ranged from 0.5636 to 0.8907 with observed heterozygosity (Hobs) ranging from 0.3333 to 0.6761 (Table 4).
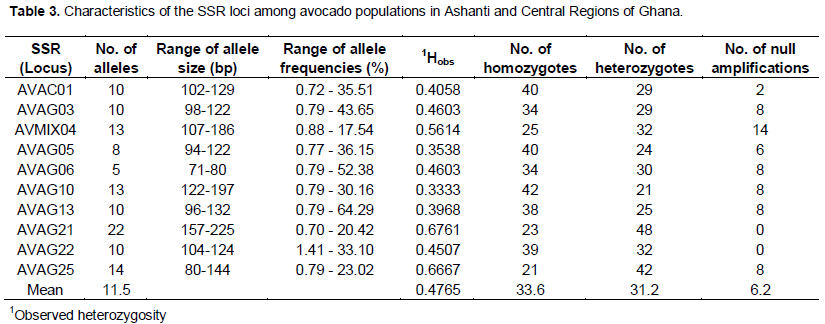
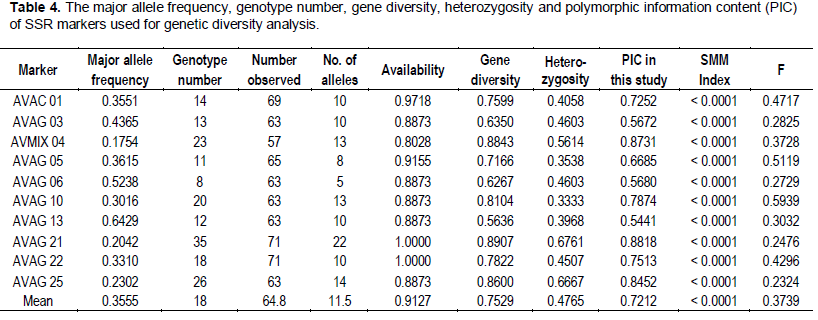
The 10 SSRs used were all highly significant (AVAC 01: c2 = 191.0, d.f. = 45, P < 0.001; AVAG 03: c2 = 166.9, d.f. = 45, P < 0.001; AVMIX 04: c2 = 196.9, d.f. = 78, P < 0.001; AVAG 05: c2 = 88.7, d.f. = 28, P < 0.001; AVAG 06: c2 = 81.8, d.f. = 10, P < 0.001; AVAG 10: c2 = 339.3, d.f. = 78, P < 0.001; AVAG 13: c2 262.8, d.f. = 45, P <0.001; AVAG 21: c2 = 468.6, d.f. = 231, P < 0.001; AVAG 22: c2 = 198.3, d.f. = 55, P < 0.001; AVAG 25: c2 = 156.9, d.f. = 91, P < 0.001). The polymorphism information content (PIC) value ranged from 0.5441 to 0.8818 with a mean of 0.7212. The proportion of loci with PIC value from 0.5 and above was 100%. We found that 70% of the accessions had PIC values of 0.6 and beyond. The Stepwise Mutation Model Index (SMMIndex) was highly significant, with mean f value of 0.3739.
Phylogenetic analyses
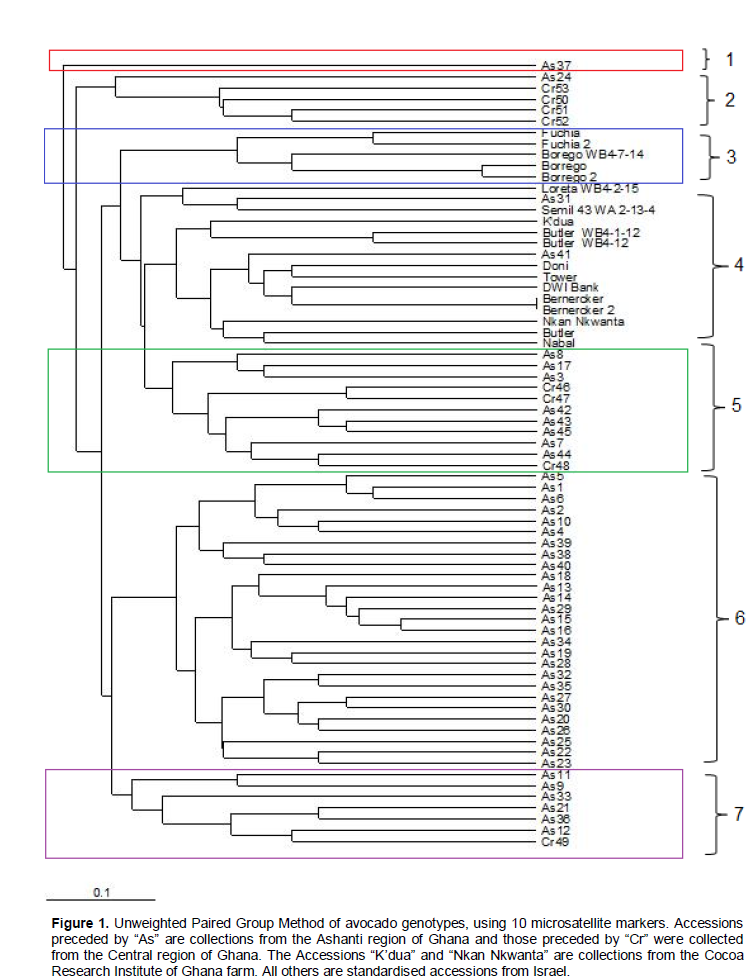
The set of markers used uniquely classified 71 individual plants in this study; it also illustrated the considerable genetic diversity that was present. The resultant dendrogram (Figure 1) defined seven distinct groups. The most genetically distinct genotype was As 37; this did not cluster with any other accession. The largest group consisted of 27 genotypes. The second distinct group had Cr 49 and As 11 at the ends.
DISCUSSION
SSR polymorphism and genetic diversity
Forty percent (40%) of the alleles scored fall outside the range of sizes previously scored and varied by one base pair more than the ones previously confirmed (Schnell et al., 2003). A high level of polymorphism was obtained in most of the loci studied, since 8 of the 10 loci revealed 10 or more alleles in the accessions analysed. The features of the locus of AVAG 21 were not much different from that of an earlier work in which it was reported to be one of the most polymorphic loci after genotyping 258 accessions (Schnell et al., 2003). The average of 11.5 allele / locus in this study was similar to the 10.4 alleles / locus reported by Ashworth and Clegg (2003), where 25 SSRs loci and 180 genotypes were used. However, the similarity obtained in this study is significantly higher than the 37 genotypes obtained in another study (Alcaraz and Hormaza, 2007). This implies that, there is a greater diversity between the avocados studies. In Schnell et al. (2003), 256 alleles were identified from the 14 SSRs loci used, ranging from 8 to 30 per locus and an average of 18.8 from the 14 SSRs was obtained. These results confirmed that the SSRs are highly polymorphic. The difference observed between Schnell et al. (2003) and the present study might be due to differences in the number of the accessions used. The observed heterozy-gosity (Hobs) value calculated from our study ranged from 0.333 in AVAG 10, to 0.6761 in AVAG 21. The average Hobs was 0.4765 which is lower than the 0.64 obtained by Schnell et al. (2003) indicating a narrower genetic diversity for the populations analysed. Unique alleles were identified within some samples (Table 2), but their frequencies were too low to allow any meaningful inferences.
Polymorphic information contents (PIC) has been found to be comparable between SSRs RFLPs and Amplified Fragment Length Polymorphisms (AFLPs) or even higher PIC value for SSRs (Smith et al., 1997; Pejie et al., 1998; Menz et al., 2004). The mean PIC and mean number of alleles obtained in our study confirms the findings of Smith et al. (1997), Pejie et al. (1998) and Menz et al. (2004). Almost all the exotic hybrids from CRIG clustered with others from the same parental line. The parental lines of avocado are the Mexican, West Indian, and Guatemalan varieties or hybrids of any two of the three varieties. The introduced hybrids from the CRIG avocado farm used in this study were predominantly of West Indian origin with one (Nabal) from a Guatemalan parent and a few inter hybrid varieties. Some of the accessions from the study area clustered with some of the varieties from CRIG (Figure 1). K’dua was found to be genetically similar to Butler WB4-1-12, Butler WB4-12, DWI Bank and Doni from West Indian parents and developed in the USA. Nabal, a Guatemalan hybrid, was also highly related to the Nkrankwanta variety, As 8 and As 17 from Ghana. This indicates that there are some plants in the study area which are West Indian and Guatemalan hybrids, thus suggesting that there might be some West Indian and Guatemalan hybrids in Ghana as previously indicated by Taah et al. (2003). Loreta WB4-2-15 was also found to be similar to As 31, whereas As 41 shared similar alleles with Doni, Tower and DWI Bank. These developments suggest that there are a number of unknown accessions of avocados in the study area which should be further studied and analysed. The phylogenetic tree suggests a wide genetic variation among the accessions genotyped. The phylogenetic tree showed highly variable levels of genetic differences between their groups.
The genetic diversity analysis shows that the SSRs used were highly polymorphic in structure. There is a wide range of diversity between the accessions which might have resulted from cross pollination and genetic mutations. Early missionaries to Ghana might have introduced mostly West Indian varieties into the country. However, there are a few Guatemalan hybrids and some hybrid varieties of the West Indian varieties in all areas where plant materials were taken for this study. There is a high probability that, seeds of the same accessions might have been used as propagules by migratory farmers and that the accessions in the Ashanti and Central regions of Ghana might be the same as the ones introduced over a century ago to Ghana by early missionaries.
CONCLUSION
In conclusion, our study provides insights into the molecular characterisation of avocado in the Ashanti and Central regions of Ghana, paving the way for further molecular genetic investigations to determine the specific avocado accessions found in Ghana.
CONFLICT OF INTERESTS
The author(s) have not declared any conflict of interest.
ACKNOWLEDGEMENTS
We are grateful to Kirkhouse Trust, UK for financially supporting this research. We are also grateful to the staff at the Biotechnology Laboratory of Cocoa Research Institute of Ghana for all their support.
REFERENCES
| Acheampong AK, Akromah R, Ofori FA, Takrama JF, Saada D, Bitton I, Lavi U (2008). Genetic characterization of Ghanaian avocados using microsatellite markers. J. Amer. Soc. Hortic. Sci. 133(6):801-809. | ||||
|
Alcaraz ML, Hormaza JI (2007). Molecular characterization and genetic diversity in an avocado collection of cultivars and local Spanish genotypes using SSRs. Hereditas 144:244-253. Crossref |
||||
|
Aldrich J, Cullis CA (1993). RAPD analysis in flax: optimization of field and reproducibility using Klen Tag 1 DNA polymerase, chelex 100 and gel purification of genomic DNA. Plant Mol. Biol. Rep. 11:128-141. Crossref |
||||
|
Ashworth VETM, Clegg MT (2003). Microsatellite markers in avocado (Persea Americana Mill.): genealogical relationships among cultivated avocado genotypes. J. Hered. 94:407-415. Crossref |
||||
|
Bassam J, Caetano-Anolles G, Gresshoff PM (1991). Fast and sensitive silver staining of DNA in polyacrylamide gels. Anal. Biochem. 196:80-83. Crossref |
||||
| Bergh BO (1969). Avocado. In: Ferwerda, F. P. and Wit, F. (Eds.), Outlines of perennial crop breeding in the tropics. Landbonwhogeschool, Wageningen, Netherlands. pp. 23-51. | ||||
| Bergh BO (1995). Avocado. In Smartt, J. and Simmonds, N. W. (Eds.), Evolution of crop plants. Harlow: Longman Scientific and Technical. pp. 240-245. | ||||
| Bergh BO, Lahav E (1996). Avocados. In Janick, J. and Moore, J. N. (Eds.), Fruit Breeding, 1: Tree and Tropical Fruits. New York: John Wiley and Sons. pp. 113-166. | ||||
|
Bergh BO, Scora RW, Storey WB (1973). A comparison of leaf terpenes in Persea subgenus Persea. Bot. Gaz. 134:130-134. Crossref |
||||
| Campbell CW, Malo SE (1976). A survey of avocado cultivars. In: Sauls, J. W., Phillips, R. L. and Jackson, L. K. (eds.). The avocado. Proc. 1st Sub-Tropical Fruit Short Course. Univ. Florida Coop. Ext. Serv. pp. 20-24. | ||||
| Felsenstein J (1989). PHYLIP - Phylogenetic inference package, version 3.2. Cladistics 5: 164-166. | ||||
| Food and Agricultural Organisation Statistical Database (FAOSTAT Database) (2001). Food and Agricultural Organisation, United Nation, Rome. | ||||
| Furnier GH, Cummings MP, Clegg MT (1990). Evolution of the avocados as revealed by DNA-restriction site variation. J. Hered. 81:183-188. | ||||
|
Goldstein DB, Linares AR, Cavalli-Sforza LL, Feldman M (1995). Genetic absolute dating based on microsatellites and the origin of modern humans. PNAS 92:6723-6727. Crossref |
||||
|
Goldstein DB, Pollock PD (1997). A review of mutation processes and methods of phylogenetic inference. J. Hered. 88:335-342. Crossref |
||||
| Gutiérrez-Diez A, Martinez-de la Cerda J, Garcia-Zambrano EA, Iracheta-Donjuan L, Ocampo-Morales JD, Cerda-Hurtado IM (2009). Study of genetic diversity of native avocado in León, México. Rev. Fitotec. Mex. 32(1):9-18. | ||||
|
Jarne P, Lagoda JL (1996). Microsatellites-from molecules to populations and back. Trends Ecol. Evol. 11: 424-429. Crossref |
||||
|
Knight RJ Jr. (2002). History, distribution and uses. In: Whiley A. W., Schaffer, B. and Wolstenholme, B. N. (Eds.), The avocado: botany, production and uses. UK: CAB International, city. pp. 1-14. Crossref |
||||
| Lavi U, Hillel J, Vainstein A, Lahav E, Sharon D (1991). Application of DNA fingerprints for identification and genetic analysis of avocado. JASHS 116: 1078-1081. | ||||
| Liu K, Muse S (2004). Power Marker: new genetic data analysis software, version 3.25. | ||||
|
Menz M, Klein RR, Unruh NC, Rooney WL, Klein PE, Mullet JE (2004). Genetic diversity of public inbreds of sorghum determined by mapped AFLP and SSR markers. Crop Sci. 44: 1236-1244. Crossref |
||||
| Mhameed S, Sharon D, Hillel J, Lahav E, Kaufman D, Lavi U (1996). Level of heterozygosity and mode of inheritance of variable number of tandem repeat loci in avocado. JASHS 121:778-782. | ||||
|
Mhameed S, Sharon D, Kaufman D, Lahav E, Hillel J, Degani C, Lavi U (1997). Genetic relationships within avocado (Persea americana Mill) cultivars and between Persea species. Theor. Appl. Genet. 94:279-286. Crossref |
||||
| Morton JF (1987). Fruits of warm climates. Julia F. Morton, Miami, Florida, 505 pp. | ||||
|
Nei M (1973). Analysis of gene diversity in subdivided populations. PNAS 70: 3321-3323. Crossref |
||||
|
Pejie I, Ajmone-Marsan P, Morgante M, Kozumplick V, Castiglioni P, Taramino G, Motto M (1998). Comparative analysis of genetic similarity among maize inbred lines detected by RFLPs, RAPDs, SSRs, and AFLPs. Theor. Appl. Genet. 97: 1248-1255. Crossref |
||||
|
Queller DC, Strassmann JE, Hughes CR (1993). Microsatellites and kinship. Trends Ecol. Evol. 8: 285-288. Crossref |
||||
| Rhodes AM, Malo SE, Campbell CW, Carmer SG (1971). A numerical taxonomic study of the avocado (Persea americana Mill.). JASHS 96:391-395. | ||||
| Röder MS, Plaschke J, Konig SU Börner A, Sorrells ME, Tanksley SD, Ganal MW (1995). Abundance, variability and chromosomal location of microsatellites in wheat. Theor. Appl. Genet. 246: 327-333. | ||||
| Saitou N, Nei M (1987). The Neighbour-joining method: A new method for constructing phylogenetic trees. Mol. Biol. Evol. 4: 406-425. | ||||
| Schnell RJ, Brown JS, Olano CT, Power EJ, Krol CA (2003). Evaluation of avocado germplasm using microsatellite markers. JASHS 128: 881- 889. | ||||
|
Sharon D, Cregan PB, Mhameed S, Kusharska K, Hillel J, Lahav E, Lavi U (1997). An integrated genetic linkage map of avocado. Theor. Appl. Genet. 95: 911-921. Crossref |
||||
|
Smith JSC, Chin ECL, Shu H Smith OS, Wall SJ, Senior ML, Mitchel SE, Kresovich S, Ziegle J (1997). An evaluation of the utility of SSR loci as molecular markers in maize (Zea mays L.): comparisons with data from RFLPs and pedigree. Theor. Appl. Genet. 95:163-173. Crossref |
||||
| Sneath PHA, Sokal RR (1973). Numerical Taxonomy. Freeman, San Francisco. 573pp. | ||||
|
Sunnucks P (2000). Efficient genetic markers for population biology. Trends Ecol. Evol. 15:199-203. Crossref |
||||
| Taah KJ, Alderson PG, Power JB (2003). Molecular approaches for the characterization of Ghanaian avocado pear (Persea americana Mill.) germplasm. Proceedings: V World Avocado Congress (Actas V Congreso Mundial del Aguacate), 19-24th October 2003 Granda Malaga, Spain. | ||||
|
Taramino G, Tarchini R, Ferrario S, Lee M, Pè ME (1997). Characterization and mapping of simple sequence repeats (SSRs) in Sorghum bicolor. Theor. Appl. Genet. 95: 66-72. Crossref |
||||
| Uptmoor R, Wenzel W, Friedt W, Donaldson G, Ayisi K, Ordon F (2003). Comparative analysis on the genetic relatedness of Sorghum bicolor accession from Southern African RAPDs, AFLPs and SSRs. Theor. Appl. Genet. 106:1316-1325. | ||||
| Verheij EWM, Coronel RE (Eds.) (1991). Edible fruits and nuts. Plant Research of South-East Asia. 2, Wagneningen, The Netherlands. | ||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


