Full Length Research Paper

ABSTRACT
INTRODUCTION
Cedrus libani A. Richard (Pinaceae) is a majestic tree of 15 to 40 m height at maturity with innumerable scriptural and historical references. It is by far the most famous and treasured tree in Lebanon. Historically, it was mentioned in mythology and cited 75 times in the Holy Bible and is used as symbol of strength and stability. References to the Cedars of Lebanon go as far back as the beginning of written script and the first temple of Solomon was built with it. In earlier times, the Cedar of Lebanon covered a large area of Lebanon’s mountains. Thus, the cedar became Lebanon’s national emblem and it is seen on the Lebanese flag.
In Lebanon, C. libani is an important forest tree species between 1000 – 2300 m and most abundant on North-facing slopes, where the impact of radiation is less severe, but in wetter locations it grows equally well on the mountain sides exposed to the prevailing rain-bringing winds (Khuri et al., 2000). Twelve (12) separate regions are known to harbor the Cedar in Lebanon: Jabal Qammoua forest, Wadi Jahannam in the Akkar area, Ehden, Bcharre, Tannourine-Hadeth, Jeij in the Jbail Mountains, and in the Jabal el-Barouk forests of the Chouf mountains, Ain Zhalta/Bmohrain, Barouk, and Maasser el-Chouf.
Cephalcia tannourinensis (Hymenoptera: Pamphiliidae), the cedar web-spinning sawfly was described as a new species by Chevin (2002). Between 1990 and 1999, the intensity had been increasing in Tannourine-Hadath El-Jebbeh cedar forest located in northern Lebanon and has caused intense defoliation of over 600 ha.
The larvae feed on cedar needles and after the last molt, they drop from the crown to the ground where they hibernate (Nemer et al., 2005). The larval diapause can last for more than one year; and a five year diapause was recorded for some species within the same genus (Gruppe, 1996). Aerial spraying with the insect growth regulator diflubenzuron was carried out in 1999-2004 resulting in considerable suppression of the cedar web-spinning sawfly population (Nemer and Nasr, 2004) but for ecological reasons development of more environmentally friendly techniques to manage populations are required (Nemer et al., 2007; Abdo et al., 2008). The danger of the spread of this insect to other cedar forests in Lebanon and the Mediterranean region led to the initiation of a UNEP/GEF project entitled “Integrated Management of Cedar Forests in Lebanon in Cooperation with Other Mediterranean Countries” that aims to develop an action plan for integrated management of forests including assessment of insects in cedar forest in the Mediterranean region with particular emphasis on the Tannourine-Hadath El-Jebbeh cedar forest. The causes of Cephalcia outbreak in the forest of Tannourine-Hadath El-Jebbeh are still not known. A variety of factors may have contributed to the population increase of C. tannourinensis during the last decade. Climatic factors and other characteristics related to the microhabitat of the forest ecosystem are thought to be among the principle causes of this outbreak. Subsequently, it was decided to initiate the present study with the following objectives: (i) determine the relationship between soil properties and survival of the prepupae of C. tannourinensis larvae as well as the annual and multi annual diapause development of this insect and (ii) understand the relation between soil temperatures and soil moisture vis-à-vis the development of the prepupae of C. tannourinensis.
MATERIALS AND METHODS
Study site
The field studies were conducted in the Tannourine cedar forest. The cedar forest of Tannourine Hadath El-Jebbeh is located in North Lebanon about 85 Km from Beirut. It falls between 34º12’ and 34°15’ of Latitude N and between 35°54’ and 35º56’ of Longitude W from Greenwich. The altitude varies between 1,430 to 1,850 m above sea level. The forest is physiologically positioned on a moderately steep sloping valley composed mostly of alluvial deposits of calcareous and basaltic deposits. The forest area is located in the supramediterranean region with an annual rainfall varying between 1,200 to 1,400 mm Food and Agriculture Organization, 2003).
Laboratory studies
Effect of different soils on the development of prepupae of C. tannourinensis
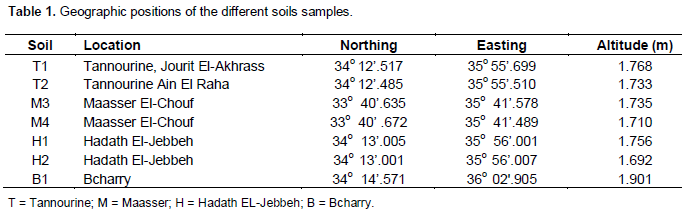
Soil sampling and analysis: Collection of soil samples and methods of analysis were done according to Bashour and Sayegh (2001). Composite soil samples were collected from Tannourine cedar forest, Maasser El-Chouf, Hadath El-Jebbeh and Bcharry (Table 1) during the first week of July 2007, which coincided to the period where mature larvae of C. tannourinensis dropping to the ground and penetrating the soil. Approximately 2 kg of soil were taken from each sampling site at a depth of 0 to 15 cm using a hand pulled auger. Soil clods were broken down and the content was mixed well. The composite sample was transferred to a clean plastic bag after being labeled. The bags were closed tightly and transported to the laboratory where each sample of soil was spread to air dry on a clean paper sheet at room temperature. Soil aggregates were also broken down by hand using gloves to accelerate the drying process. After 10 days each sample was crushed and ground to particles of uniform size using porcelain crushers and sieved through a 2-mm sieve. Then each sample was stored properly in a well labeled and air tight clean plastic container for further analysis.
(i) Moisture content: Before air drying the samples, an amount of approximately 200 g of fresh soil was taken from each bag and transferred to a weighed metal can with a tight fitting lid; the weight of the can and the soil was taken using a mettler balance. The cans were placed in a drying oven at 110°C for a period of 24 h, cooled in a dessicator and weighed again. Moisture content was calculated according to the following formula:
Weight of water = Weight of air dried soil – Weight of oven dried soil
% Moisture content = (Weight of Water / Weight of oven dried soil) x 100
(ii) Organic matter content (combustion method): The oven dried soil samples described above were placed in a furnace at 350°C for four hours. The cans were then closed tightly and transferred into desiccators in order to cool; then were weighed again. Organic matter content was calculated on oven dried basis (Bashour and Sayegh, 2001).
(iii) Soil texture: The soil particle size distribution was determined by the Bouyoucos Hydrometer Method. Fifty grams of oven dried fine texture soil were taken from each sample and then transferred into a baffled stirring cup; and was filled with distilled water to half full, 15 ml of sodium hexametaphosphate (1N) was also added. Then the mixture was stirred in an electrically driven mixer for 10 min. The stirred mixture was transferred to a settling cylinder, the soil suspension was shaken vigorously in a back and forth manner. After 20 s, the hydrometer was inserted into the cylinder and the readings were taken after 40 s. This reading indicated the percentage of clay and silt particles in the soil suspension. A second reading was taken after 2 h for the determination of clay. Temperature was measured and corrections were done according to the outlined procedure in (Bashour and Sayegh, 2001).
(iv) Exchangeable cations: The exchangeable calcium (Ca), sodium (Na), potassium (K) and magnesium (Mg) contents were measured by taking 3 g of air dried soil from each sample and transferring them into a plastic falcon tube. A volume of 30 ml 1.0 N ammonium acetate were added to each sample and the mixture was shaken on an orbital shaker for 30 min, then all the samples were filtered and the filtrate were stored in a refrigerator at 4°C for further analysis. Ca, Na and K were measured using an AFP 100 Flame photometer, (Biotech Engineering management Co. LTD, U.S.A.) and Mg was measured using an AA 6300 Atomic absorbance spectrophotometer (SHIMADZU, Japan) (Bashour and Sayegh, 2001).
(v) Micronutrients (Fe, Zn, Mn and Cu) analysis: Available micronutrients in soil (iron, zinc, manganese and copper) were extracted by taking about 5 g of air dried soil were taken from each sample and transferred to a plastic falcon tube. A volume of 20 ml of diethylenetriaminepentaacetic acid (DTPA) solution was added to each tube. The tubes were then shaken on an orbital shaker for a period of 30 min. and filtered using a Whatman filter paper No. 40 in order to be measured on an AA 6300 Atomic absorbance spectrophotometer (SHIMADZU, Japan) (Bashour and Sayegh, 2001).
(vi) Available phosphorus (P): The determination of available phosphorus in the soils was done by placing 3 g of soil from each air dried soil sample into a 100 ml falcon tubes. 50 ml of sodium bicarbonate (0.5 M) were added to each tube and placed on the shaker for 30 min. The mixtures were then filtered using a Whatman filter paper No. 40 and a clear solution was attained. Eight drops of Sulfuric acid (H2SO4 2.5 M) were added to lower soil pH. Molybdate solution and 8 ml ascorbic acid were added, swirled and washed with distilled water in order to remove the C02 gas and form a complex blue color. Phosphorus was measured on an Aquamate spectrophotometer (Thermo Electron Corporation, USA) (Bashour and Sayegh, 2001).
(vii) Free Calcium Carbonate (CaCO3) Content: 3 g from soil sample were placed in 250 ml beaker, 30 ml of standardized (1.0 N) hydrochloric acid (HCL) were added. The beakers were each covered by a watch glass and boiled for 5 min then cooled down and filtered using a Whatman filter paper No. 40. Phenolphthalein indicator was added to the excess acid. The filtrates were measured by back titration with 0.5 N sodium hydroxide (NaOH) (Bashour and Sayegh, 2001).
(viii) pH and electric conductivity (EC): A total weight of 10 g of each air dried soil sample were transferred into a 50 ml falcon tube, 25 ml of distilled water were added to each tube and then shaken on an electric shaker for 30 min. The mixtures were then filtered using a Whatman filter paper No. 41 and the pH values were measured by a Thermo Orion pH meter model 410 A +. For comparison to a peat moss sample (Russian made), a saturation paste was made of 10 g. of peat and 70 ml of distilled water in a covered beaker for a period of 24 h. Afterwards, the paste was filtered on a Whatman filter paper No. 41 and the filtrate was transferred to a clean 200 ml falcon tube. EC was measured using a Mettler Toledo, M226 Conductivity meter. The readings were preformed under constant laboratory temperature at 24°C (Bashour and Sayegh, 2001).
(ix) Cation exchange capacity (CEC): About 1 g were taken from each soil sample and transferred to a 200 ml falcon tube to which a volume of 8 ml 1 N Sodium acetate (NaOAC) was added. The tubes were shaken for 5 min on an electric shaker. Then the caps and the sides of the tubes were washed well with NaOAC solution. The clean tubes were centrifuged for 3 min. at a speed of 2000 rpm in a Durafuge 200 centrifuge (Precision, France). The supernatant was discarded. This operation was repeated three times. Afterwards, the samples were washed four times with 8 ml ethanol and centrifuged. The EC of the supernatant became 40 microseimens per cm. or even less. Then 8 ml of ammonium acetate (NH4OAC) were added to each sample and centrifuged for 3 min, the supernatants were collected and placed into a 50 ml volumetric flask. Sodium (Na) was measured directly on an AFP 100 Flame photometer (Biotech Engineering Management Co. LTD, USA) (Bashour and Sayegh, 2001).
Experimental setup
Cedar branches infested with second instar larvae of C. tannourinensis were collected from Tannourine forest during the month of June 2007 and brought to the laboratory where they were placed in an incubator set at 20°C with a photoperiod of 14:10 (L:D). The larvae were allowed to mature under these conditions. Prepupae (mature larvae) were collected once they ceased feeding and dropped from cedar branches. The prepupae were then transferred to the different soil types (Table 1) in addition to two reference soil types namely peat moss (Kitexim, Kaliningrad, Russia) and pure sand. A total weight of 40 g of soil was taken from each soil type with the exception of peat moss where only 20 g were taken. The soils were placed in 50 ml glass beakers. The prepupae were placed on the top of soil and allowed to penetrate freely in it. The beakers were then sealed with parafilm in order to reduce evaporation and thus maintain a relatively constant humidity. A total number of six prepupae were placed in each beaker and each soil type was replicated three times. The total number of soil types and origin was nine. The beakers were placed in the incubator under a constant temperature of 20°C equal to the average soil temperature observed in the forest of Tannourine during the months of June and July.
Data on prepupal mortality, number of prepupae that developed into pronymphs (complete eye development) and those into eonymphs (absence of eye formation) were taken after 105 days. This period is necessary for complete eye formation based on field observations at the Tannourine cedar forest.
Statistical analysis
The experiment was a completely randomized design with one factor, the soil origin and types. ANOVA 1 was performed and means were separated by LSD. Correlation and linear regression analysis were performed between the different soil factors (moisture, nutrients, texture, etc.) and the larval mortality and the formation of eye.
Effect of soil temperature on the development of Prepupae of C. tannourinensis
Soil sampling
In this experiment, soils from Tannourine forest only were used since prepupae of C. tannourinensis are known to live in these soils. Soil samples from a previously outbreak area in the Tannourine forest were collected in sealed plastic bags in July 2007 and directly used in the experiment to reduce humidity loss.
Experimental setup
The experiment was set as described previously in (soil sampling and analysis/experimental setup) and each beaker was sealed with parafilm after the prepupae were placed on top of the soil. Treatments were replicated three times and 20 prepupae were used per treatment. Three replicates of each treatment were placed in an incubator held at 20°C and another three replicates were placed in an incubator held at 10°C. Data on larval mortality, number of prepupae that developed into pronymphs (complete eye development) and those into eonymphs (absence of eye formation) were taken after 105 days.
Statistical analysis
The number of dead prepupae, pronymphs and eonymphs were analyzed by t-test by comparing the means of both temperature regimes.
Effect of soil moisture on the development of Prepupae of C. tannourinensis
Soil sampling
Soil samples from a previous outbreak area in the Tannourine forest were collected in July 2007. Soil moisture was adjusted to three levels: 5, 20 and 40%. The soil moisture of the samples was determined following the method described in experiment (soil sampling and analysis). The adjustments of moisture were done by either adding distilled water to the sample to reach the level required or oven drying the samples and regulate the measurement to reach the required moisture level.
Experimental setup
The experiment was set as described in experiment (effect of soil temperature on the development of Prepupae of C. tannourinensis / soil sampling) and each beaker was sealed with parafilm after the prepupae were placed on top of the soil. Soil samples were replicated three times with 10 prepupae in each for each moisture level: 5, 20 and 40%. The sealed beakers were placed in an incubator held at 20°C. Data on larval mortality, number of larvae that developed into pronymphs (complete eye development) and those into eonymphs (absence of eye formation) were taken after 105 days.
Statistical analysis
The design of the experiment was set as a completely randomized design (CRD) with one factor, soil moisture. ANOVA 1 was performed and means were separated by LSD. Linear and nonlinear curves were determined to evaluate the relationship between moisture levels and eonymphs.
RESULTS AND DISCUSSION
Effect of different soils on the development of Prepupae of C. tannourinensis
The main purpose of this experiment was to study the effect of different soil physical and chemical properties and their influence on the development of the C. tannourinensis prepupae conducted under stable laboratory conditions (temperature, light and moisture).
Soil origins and analysis
Moisture content
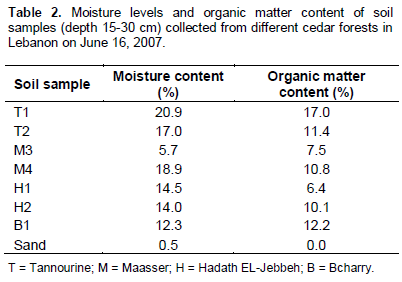
Soil samples were collected within two days during the second week of June 2007. Soil moisture measurements showed an elevated moisture percentage in the samples from Tannourine forest (20.9 and 17.0%) followed by one location in Maasser El-Chouf (M4) (18.9%). The moisture percentage in H1 and H2 from Hadath El-Jebbeh forest were 14 and 14.5 %, respectively (Table 3). Soil moisture content depended on the season and the annual precipitation of a specific region.
Organic matter content
Soil organic matter varies between 1 and 5% in most Lebanese arid and semi arid regions. This was not the case in our soil samples due to the fact that the samples were taken from cedar forests. The dense canopy of these forests affected the soil organic matter and raises the percentage to levels ranging between 6.4 and 17% (Table 2).
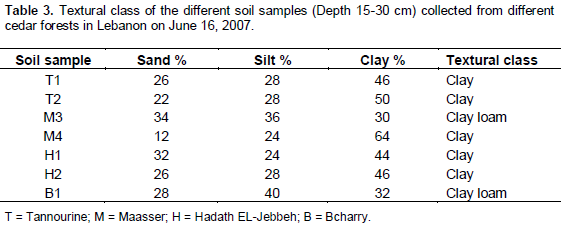
Soil texture
The results of soil particle size distribution showed that the soil samples from different locations are of clayey type and can be further classified into Red Mediterranean soil (Table 3). The clayey Mediterranean soil is characterized by complex and diverse parental material with various drainage conditions and seasonal moisture fluctuation. The soil samples can be further classified into Reddish Brown and Red Ferrallitic soils and they were first described in 1930 in many Mediterranean countries under the name of Terra-Rossa (Marcelin, 1942; Bashour and Sayegh, 2001).
Exchangeable cations (Ca, Na, K, and Mg) analysis
The results for exchangeable cations are presented in table 4. Results indicate that the Tannourine soil is very rich in potassium (K); because available K which was extracted with NH4OAC was higher than 500 mg/kg. This is due to the mineral types and texture of the soil which is clayey. Clayey soils prohibit the fast leaching of K which originates from the fresh plant and tree residues. K is very leachable in humus or well decomposed plant material. Similar levels of K were obtained in Hadath-El- Jebbeh, Bcharry and one location of Maasser El-Chouf, where K was higher than 300 mg/kg, except for one sample M4 from the Maasser El-Chouf cedar forest. The low K level of M4 sample may be due to the high rainfall, snow rates and high leaching in addition to the lack of vegetation cover to protect soil from strong wind which prevails in the area which blows away the fine mineral soil particles.
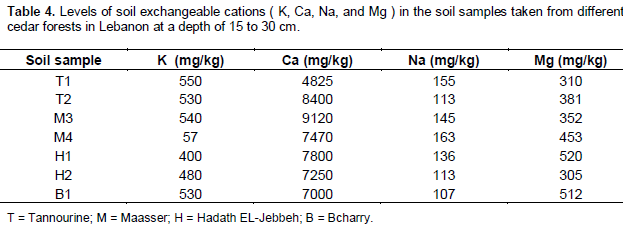
Calcium (Ca) levels were higher than 3500 mg/kg in all soil samples (Table 4); this is considered very high according to Bashour and Sayegh (2001). The levels of sodium (Na) in all the samples was medium/low (Table 4) according to Bashour and Sayegh (2001). Soil Na ranged between 100 and 150 mg/kg. Unlike other areas in Lebanon, the Tannourine cedar forest samples belong to a wild uncultivated soil having undisturbed profiles with good infiltration levels, therefore salinity levels are way below the damaging threshold and salt content in soils do not cause problems for plant growth. Results for magnesium (Mg) indicated that its level in the soil samples varied between high to very high (300 and 500 mg/kg) Bashour and Sayegh (2001) (Table 4).
Micronutrients (Fe, Zn, Mn and Cu) analysis
The soil micronutrients are essential for plant growth and are needed in minute quantities. They originate from weathered minerals, are more soluble in acidic solutions or acidic soils, are directly influenced by high pH, and they are usually low in soils of arid regions especially calcareous soils.
Results showed that the soil samples were very high in iron (>10mg/kg) Bashour and Sayegh (2001) (Table 5). This may be due to the nature of parent rocks and the high levels of humus and organic matter in the soil. Also the results of available Mn, and Cu were high in the soil. As for the available Zinc (Zn), its level was found to be medium in all the samples, ranging between 1.2 to 4 mg/kg.
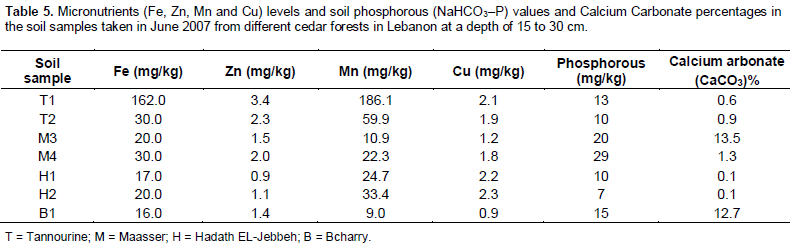
Available phosphorus (P)
Results of available phosphorus revealed that Maasser El-Chouf soils have the highest levels followed by Bcharry with 15 mg/kg, then Tannourine with 13 and 10 mg/kg in both samples and finally by Hadath El-Jebbeh (7-10 mg/kg) the least amount of available phosphorous which is considered sufficient for forest growth (Table 5).
Free calcium carbonate (CaCO3) content
Results of calcium carbonate analysis (Table 5) indicated that Tannourine soils are very low in CaCO3 according to the recommended ranges Bashour and Sayegh (2001). Results of analysis indicated that samples (M3 and M4) collected from Maasser El-Chouf were high in CaCO3 levels. The other five samples collected from Bcharry and Hadeth El-Jebbeh forests were medium in CaCO3 content. Calcium carbonate affects the moisture characteristics of soils. It can help in the formation of soil aggregates and interferes in its particle size distribution. Moreover, CaCO3 can be considered as a soil cementing agent helping or improving the formation of soil aggregates.
pH and electric conductivity (EC)
Soil pH of Lebanese cedar forests was found to be slightly basic with minute differences between the different forests (Table 6). Electrical conductivity (EC) is used to determine the total concentration of solutes and therefore reflects the salinity levels of soil; it has a critical effect on the germination and emergence stages of seedlings. Results showed that soils of different cedar forests have low EC as it is the case in all Lebanese soils.
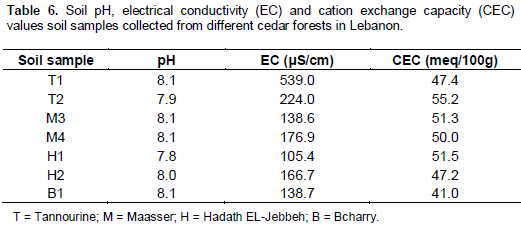
Cation exchange capacity (CEC)
CEC is defined as the number of cation adsorption sites per unit weight of soil or the total sum of exchangeable cations that a soil can adsorb. CEC is expressed in centimole per kilogram (cmole/kg) of oven dried soil. CEC values were found to be high in all forest soil samples with minor differences between them (Table 6). A high CEC value (>25 meq/100 g) is a good indicator for a soil rich in clay and organic matter content, therefore it will hold high amounts of cations and reduce leaching Bashour and Sayegh (2001).
Laboratory experiment with Prepupae of Cephalcia tannourinensis
Prepupa mortality was significantly affected by soil types (ANOVA; F =10.827; df = 8, 26; P<0.0001) (Table 7). C. tannourinensis prepupa survived well in soils coming from Tannourine (T1 and T2), where the maximum mortality was 27.8%. Prepupae have constructed an earth cell chamber and thus protected themselves from physical and biological disturbances. The highest mortality was observed in soils originating from Maasser El-Chouf (M3) and in the two reference soils, namely peat and sand where the mortalities were 100, 77.8 and 100%, respectively. The Hadath El- Jebbeh soil (H2), the Maasser El-Chouf (M4) and the soil of Bcharry (B1) showed similar mortality rate to that of the two samples from Tannourine (T1 and T2).
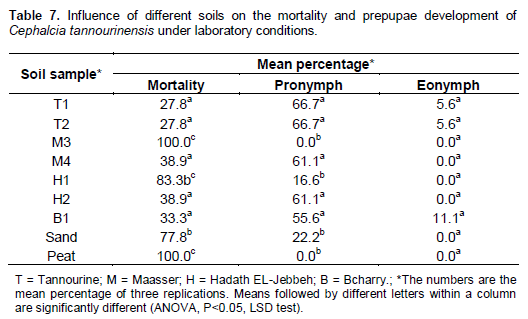
Observations showed that the high mortality in H1 was related to an attack by Beauveria fungus at the developing prepupae. Pure sand and pure peatmoss samples resulted in an elevated mortality rate due to the inability of C. tannourinensis larvae to form the earth cell chamber or loge as influenced directly by strictly reduced moisture levels in both cases.
Correlation analysis between different soil properties namely, pH, EC, CaCO3%, P, K, Ca, Mg, Na, Fe, Zn, Mn, Cu, textural class, and organic matter content versus prepupal mortality indicated that only two properties affect prepupal survival; soil moisture and textural class (Pearson correlation = - 0.782 for moisture and 0.791 fortexture). The relationship between prepupa mortality and soil moisture is inversely related. The relationship between prepupa mortality and texture was direct and mortality increased as the soil texture changed from clayey to clay loam, peat, and sand, respectively. The highest mortality was seen in soils with low moisture content; Maasser (M3), pure sand and peat, respectively (Figure 1).
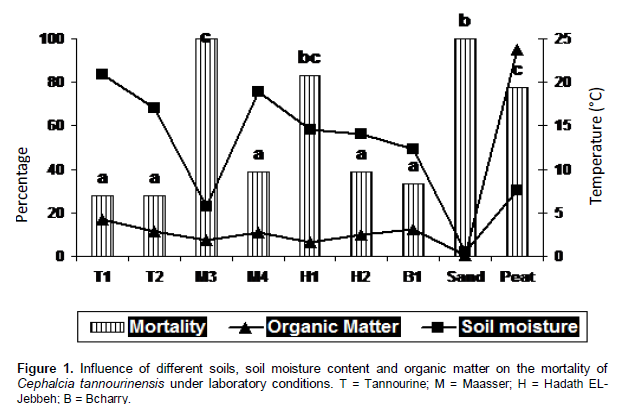
The above results demonstrated that limiting factors for the survival of C. tannourinensis prepupae are moisture and soil texture. Only few studies have suggested that moisture controls diapause development directly in arthropods (Tauber and Tauber, 1976). Some studies indicated that water has an essential role in post-diapause developmental phase of certain arthropods (Danks, 1987; Tauber et al., 1998). The European corn borer, Ostrinia nubilalis, has a two-phase diapause process, whereby changes in photoperiod terminated diapause but water was required to activate the neuroendocrine system to allow post-diapause development to be completed (Beck, 1967). Some studies conducted on tropical insects found that the presence or absence of moisture appears to play a major role in diapause initiation (Tauber and Tauber, 1976; Seymour, 1991).
The rate of prepupal development into eonymph was almost the same in all soils (ANOVA; F=0.792; df = 8. 26; P>0.05) whereas development into pronymph was significantly different between the different soils (ANOVA; F =9.630; df = 8. 26; P<0.0001). Pronymph was positively correlated with the moisture factor and texture (Pearson correlation = 0.814 for moisture and -0.812 for texture) under the conditions of this study.
All other factors showed no correlation with the development of pronymph. The relationship between pronymph and moisture was positive; as soil moisture increased from 5, 20 to 40%, respectively, the number of pronymph also increased. The relationship between the number of developing pronymph and texture was inversely related. The proportion of pronymph increased as the soil texture changed sequentially from sand, peat clay loam to clay.
Effect of soil temperature on the development of Prepupae of C. tannourinensis
Results show that the two studied temperatures that is, 10 and 20°C did not affect the mortality of C. tannourinensis under laboratory conditions (Table 8). Soil moisture was maintained constant at 18% throughout the experiment. A high number of pronymphs was obtained under a temperature of 20°C and was significantly different from that at 10°C. Eonymphs were not significantly different between the two temperature regimes although a higher number of eonymphs was recorded in the results at 10°C temperature. Battisti (1994a, b) demonstrated that prolonged diapause as an eonymph occurred in prepupae of C. arvensis just after soil penetration at temperature below 12°C. He also found that mature larvae that entered the soil at tempe-ratures above 12°C changed immediately into pronymphs and these individuals will emerge during the next spring. These findings confirm our results in that a higher number of pronymphs was obtained when mature larvae were placed in soils kept at temperature of 20°C than when placed at 10°C (Table 8).
The none significance between eonymph numbers in both temperature regimes could be attributed to an interaction between the temperature and soil moisture, since soil moisture was found to be significant with both mortality and pronymph development in the first experiment.

Effect of soil moisture on the development of Prepupae of C. tannourinensis
When exposed to different moisture regimes, physiological development of C. tannourinensis larvae showed significant differences in the number of dead, pronymps and eonymphs (Table 9). The highest number of dead larvae was recorded at 5% soil moisture level, and it was significantly higher from the 20 and 40% soil moisture regime (ANOVA, F=9.41, df=2, P<0.05). These results confirm the results of experiment 1 where a high mortality was found in the soils with low soil moisture content that is, M3, sand and peat moss.
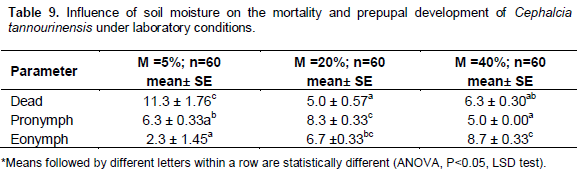
Pronymphs were significantly higher in 20% soil moisture level as compared to the other two that is,. 5 and 40% (ANOVA, F =38, df= 2, P < 0.001). These results may be attributed to the fact that the lowest moisture level led to highest mortally rate and therefore the number of pronymphs was reduced. In the high moisture level, the number of larvae entering an annual diapause cycle was low due to the fact that a high proportion of the larvae entered a multi-annual diapause cycle reflected by the high number of eonymphs recorded (8.7 ± 0.33). The second explanation may be related to the temperature of the experimental set up which was kept constant at 20°C. In the previous experiment a higher number of pronymphs was obtained at 20°C as compared to 10°C. Therefore the interaction between temperature and soil moisture may also affect the development of prepupae. Further experiments where both the temperature and soil moisture are varying are required to demonstrate the interaction effect of these two limiting factors in the development of prepupae of C. tannourinensis.
ANOVA analysis of eonymphs was significantly different between the different soil moisture levels (ANOVA, F= 13.48, df=2, P<0.01). Eonymphs numbers were highest in soils with 40% moisture content followed by 20 and 5% soil moisture content, respectively.
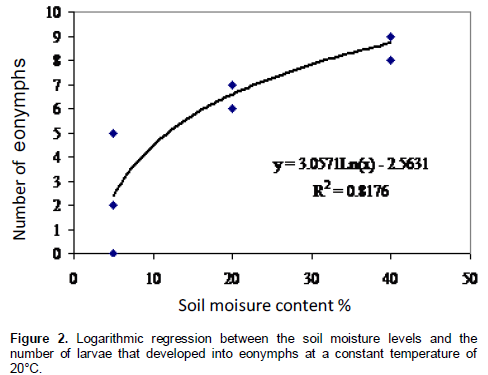
Correlation analysis between soil moisture levels and the number of dead larvae and pronymphs respectively were not significant. However, the number of eonymphs was positively correlated with moisture levels (Figure 2). Regression analysis between soil moisture levels and the number of eonymphs was significant and is defined in the following logarithmic equation:
Y= -2.56 + 3.06 Ln (x)
Where, Y represent the number of eonymphs and x the soil moisture levels.
The equation is applicable only for soil moisture contents below 40%. The experiment showed that at 40% the earth cell chamber made by the prepupae of C. tannourinensis resisted well the pressure of water.
Distribution of Prepupae in the cylinders
All pronymphs and eonymphs were present in the lower 10 cm of the plastic cylinders indicating that prepupae preferred to go deep in the soil where soil moisture content would be greater than in the upper soil layers of the cylinders. Battisti (1993) found that eonymphs are usually found deeper in the soil than pronymphs which is different from the results in the plastic cylinders. One explanation could be due to the experimental setup which is not the same used by Battisti (1993) who observed the distribution of prepupae in natural conditions.
Weight analysis of pronymphs and prepupae was significantly different between the annual individuals and the multi-annual individuals (Figure 3). The results are similar to those obtained by Battisti (1993) where biennial individuals of C. arvensis were heavier than annual individuals.
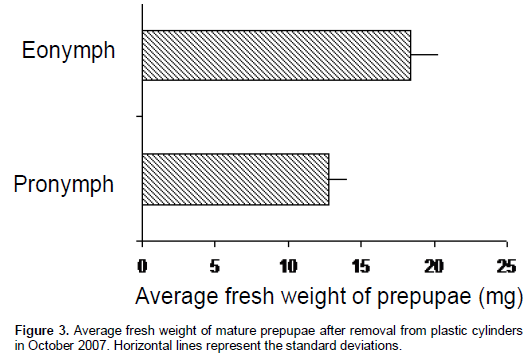
The difference observed in prepupal weight among annual and multiannual individuals could be related to the kind of larval feeding since, once in the soil, the prepupae construct a cell earth chamber and they do not feed. This may have a biochemical origin and can be correlated with climatic factors and larval development on the trees as suggested by Battisti and Cescatti (1994) and therefore the fate of prepupae would be determined at the feeding larval stages.
In this study, the diapausing cycles of C. tannourinensis were demonstrated to be related to three soil properties: soil moisture content, soil temperature and soil texture. The long life cycle and the prolonged diapause observed in C. tannourinensis is an example among 150 other insects belonging to different orders and show the pattern (Saulich, 2010). In many other species, however, the requirement for specific diapause-terminating conditions (Tauber and Tauber, 1976) is strict and/or fitness is considerably compromised without their intervening effect (Hodek, 1996 ; Hodek, 2002 ; Hodek, 2003). For instance, the maintenance phase, under constant laboratory conditions that were used for diapause induction and initiation, will continue until all larvae of the fly Chymomyza costata die, while the termination may proceed only at low temperatures (Koštál et al., 2000). In the laboratory, the maintenance and termination phases can be clearly separated in some insects by specific settings of the environmental conditions. In the field, complexity, fluctuations and linear changes of environmental conditions make the distinction more difficult.
Nevertheless, it has been shown that specific conditions/stimuli often do participate in the termination of diapause in the field, even if they are not strictly required in the laboratory. This is ecologically meaningful because the initiation of diapause may take place during very different periods of the year in different individuals of the same population. Each individual then maintains the diapause for a different time before the advent of the adversity period, which then serves as synchronizing stimulus and prevents untimely (premature) termination of diapause. Termination then takes the form of a distinct eco-physiological phase, during which diapause intensity decreases to its minimum level and subsequent resumption of direct development is enabled. Numerous examples of the effects of diapause-terminating conditions are reported in the literature. Chilling is the most common factor terminating many winter diapauses in the field (Tauber et al., 1986; Hodek, 1996; Hodek, 2002). Tanno (1970) found that freezing was a necessary factor for termination of pre-pupal diapause in the Japanese poplar sawfly Trichiocampus populi, both in the field and laboratory. Most of the summer diapauses and, perhaps, even some rare cases of winter diapauses, are terminated in the field by the change of photoperiodic signal (Masaki, 1980; Tauber et al., 1986, Ito, 1988). Contact with water was reported to serve as the terminating factor for summer diapauses of the larvae of the stem borer Busseola fusca (Okuda, 1990), and of the eggs of the chrysomelid beetle Homichloda barkeri (Nahrung and Merritt, 1999) (but see also the discussions in: Tauber et al., 1998; Hodek, 2003). In some crustaceans, drying of the sediment increases emergence from diapausing eggs upon r e-hydration (Arnott and Yan, 2002).
The results obtained in these series of experiments demonstrated that the C. tannourinensis prepupae transformation into pronymphs and eonymphs is more related to soil moisture content and temperature. Prepupae of C. tannourinensis survive well in clayey, silty and silty loam soils but cannot survive or continue their development in sandy or organic soils. Soil moisture is critical for the survival of C. tannourinenis prepupae: soils with moisture contents lower than 10% when C. tannourinensis drops to the ground and enter the soil are lethal to the prepupae. Soil moisture content and soil temperature are the most limiting factors in the determination of annual diapause individuals and multi annual diapause individuals.
Tannourine Hadath El-Jebbeh and Bcharry soils constitute favorable microhabitat environment for the development of C. tannourinensis as compared to the Chouf cedars where the soil moisture content and organic matter are lower. The presence of an organic horizon layers enhanced the survival of C. tannourinensis prepupae. In these soils the water holding capacity is greater than in mineral soils and thus soil moisture remains adequate all throughout all the summer season. However, not all the diapause was explained by these three properties. Other factors related to feeding habit of the larvae and endocrinology studies are to be exploited to determine their relation to diapause. Considerable increase of C. tannourinensis populations has been observed at Bcharry cedar forest where a high number of pronymphs was detected indicating an increase in the temperature of soil that is favorable for their development. Suppression tactics are required if these conditions will remain prevalent.
CONFLICT OF INTERESTS
The author(s) have not declared any conflict of interests.
ACKNOWLEDGEMENTS
This research was funded by a grant to the Lebanese Ministry of Environment from the Global Environment Facility and the United Nations Program (Grant Endorsement Decree No 12833 for the project “Integrated Management of Cedar Forests in Lebanon in Cooperation with Other Mediterranean countries. We thank Mr Challitah Tanios for his technical assistance with the fieldwork.
REFERENCES
|
Abdo C, Nemer N, Nemer G, Abou Jawdah Y, Atamian H, Kawar N (2008). Isolation of Beauveria species from Lebanon and evaluation of its efficacy against the cedar web-spinning sawfly, Cephalcia tannourinensis. BioControl 53:341-352. Crossref |
||||
|
Arnott SE, Yan ND (2002). The influence of drought and reacidification on zooplankton emergence from resting eggs. Ecol. Appl. 12:138-153. Crossref |
||||
| Bashour I, Sayegh A (2001). Methods of analysis for soils in arid and semi-arid regions. American university of Beirut Lebanon. | ||||
|
Battisti A (1993). Bionomics of the spruce web-spinning sawfly Cephalcia arvensis Panzer in Northeastern Italy (Hym., Pamphiliidae). J. Appl. Entomol. 115:52-61. Crossref |
||||
|
Battisti A (1994a). Voltinism and diapause in the spruce web-spinning sawfly Cephalcia arvensis. Entomol. Exp. Appl. 70:105-113. Crossref |
||||
|
Battisti A (1994b). Effects of entomopathogenic nematodes on the spruce web-spinning sawfly Cephalciaarvensis Panzer and its parasitoids in the field. Biocontrol Sci. Technol. 4(1):95-102. Crossref |
||||
| Battisti A, Cescatti A (1994). Temperature-Dependant growth model for eggs and larvae of Cephalcia arvensis (Hymenoptera: Pamphiliidae). Environ. Entomol. 23(4):805-811. | ||||
|
Beck SD (1967). Water intake and the termination of diapause in the European corn borer, Ostrinia nubilalis. J. Insect Physiol. 13:739-750. Crossref |
||||
| Chevin H (2002). Cephalcia tannourinensis n. sp., nouveau ravageur du Cèdre du Liban (Hymenoptères Pamphiliidae). L'entomologiste 58(5-6):241-242. | ||||
|
Danks HV (1987). Insect Dormancy: an Ecological Perspective. Biological Survey of Canada Monograph series no.1. National Museum of Natural Sciences, Ottawa. PMCid:PMC1148414 |
||||
| Food and Agriculture Organization (FAO) (2003).The state of the World's Forests. FAO, Rome. | ||||
|
Gruppe A (1996). Model of the importance of exogenic factors influencing the variable dormancy of prepupae of Cephalcia abietis (Hymenoptera: Symphyta: Pamphiliidae). Entomologia Generalis 21(1-2):95-105. Crossref |
||||
| Hodek I (1996). Diapause development, diapause termination and the end of diapause. Eur. J. Entomol. 93:475–487. | ||||
|
Hodek I (2002). Controversial aspects of diapause development. Eur. J. Entomol. 99:163-173. Crossref |
||||
|
Hodek I (2003). Role of water and moisture in diapause development (a review). Eur. J. Entomol. 100:22-232. Crossref |
||||
|
Ito K (1988). Diapause termination in Cletus punctiger Dallas (Heteroptera: Coreidae) in the field. Jpn. J. Appl. Entomol. Zool. 32:63-67. Crossref |
||||
|
Khuri S, Smeary MR, Baalbaki R, Maunder M, Talhouk SN (2000). Conservation of the Cedrus libani populations in Lebanon: history, current status and experimental application of somatic embryogenesis. Biodivers. Conserv. 9:1261-1273. Crossref |
||||
| Marcelin P (1942). Sur les problèmes des terres rouges. Soc. Languedocienne Georgr. 13:1-9. | ||||
|
Masaki S (1980). Summer diapause. Annu. Rev. Entomol. 25: 1–25. Crossref |
||||
|
Nahrung HF, Merritt DJ (1999). Moisture is required for the termination of egg diapause in the chrysomelid beetle, Homichloda barkeri. Entomol. Exp. Appl. 93:201-207. Crossref |
||||
| Nemer N, Demolin G, Kawar N, Kfoury L, Zakhour E (2005). Monitoring of the new cedar web-spinning sawfly, Cephalcia tannourinensis n.sp. in cedar forests of Lebanon. In: F. Lieutier and D. Ghaioule (Eds). Entomological Research in Mediterranean Forest Ecosystems, Paris, INRA Publications. pp. 247-255. | ||||
|
Nemer N, Kawar NS, Kfoury L, Frérot B (2007). Evidence of sexual attraction by pheromones in the cedar web-spinning sawfly. Can. Entomol. 139(5): 713-721. Crossref |
||||
| Nemer N, Nasr J (2004). Saving the Cedars of Lebanon. Biocontrol News and Information 25(1): 9N-11N. | ||||
|
Okuda T (1990). Significance of water contact as a factor terminating larval diapause in a stem borer, Busseola fusca. Entomol. Exp. Appl. 57: 151–155. Crossref |
||||
|
Saulich A (2010). Long life cycles in insects. Entomol. Rev. 90(9): 1127-1152. Crossref |
||||
| Seymour JE (1991). Aspects of the ecology of Microgasterdemolitor, a larval parasitoid of Heliothis armigera and Heliothis puntigera in Australia. PhD Thesis, James Cook University, Townsville. | ||||
| Tanno K (1970). Frost injury and resistance in the poplar sawfly, Trichiocampus populi Okamoto. Contributions from the Institute of Low Temperature Science Hokkaido University. B 16:1-41. | ||||
|
Tauber MJ, Tauber CA (1976). Insect seasonality: diapause maintenance, termination, and postdiapause development. Annu. Rev. Entomol. 21:81-107. Crossref |
||||
| Tauber MJ, Tauber CA, Masaki S (1986). Seasonal Adaptations of Insects. Oxford University Press, Oxford. | ||||
| Tauber MJ, Tauber CA, Nyrop JP, Villani MG (1998). Moisture, a vital but neglected factor in the seasonal ecology of insects: Hypotheses and tests of mechanisms. Environ. Entomol. 27:523-530. | ||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


