ABSTRACT
Common bean landraces represent an important source of genetic resources, which is under-exploited by Kenyan crop breeding programs. The objective of this study was to characterize 51 common bean landraces from South Western Kenya for resistance to Pythium root rot using 5 peroxidase gene (POX) markers. Following infection with Pythium spp., 11.77, 54.90 and 33.33% of the landraces were moderate-resistant, susceptible and highly susceptible, respectively. A total of 1119 alleles were amplified by the 5 primers, ranging from 3 to 8 alleles per locus. The polymorphism information content (PIC) of the POX markers varied from 0.10 to 0.47. The dendrogram generated using similarity coefficients and un-weighted pair group method with arithmetic averages (UPGMA) did not reveal any unique groupings according to their reaction to Pythium root rot disease. Population structure analysis separated the germplasm into 3 groups and all the groups contained landraces exhibiting both moderate resistance and susceptible to Pythium root rot. The lack of distinct grouping of landraces based on resistance suggests presence of significant genetic variability and different sources of resistance to Pythium root rot. These findings give valuable information for breeders and serve as a baseline for development of cultivars with Pythium root rot resistance.
Key words: genetic variability, population structure, peroxidase gene markers, Pythium root rot, resistance.
Common beans (Phaseolus vulgaris L.), are the most important grain legume, second to maize as a food crop in Kenya. Africa contributes 17% of the world’s total yield, of which 70% is from Eastern Africa. Kenya produces 400 - 1200 kg/ha, mainly from intercropping in small scale farms and an average household production of 430 kg/ha in South Western Kenya (FAOSTAT, 2013). Common beans provide a cheap source of protein and minerals (iron and zinc) to humans. In addition, consumption of bean grains provides humans with various health benefits including reduction of cholesterol level and coronary heart diseases (Mattei et al., 2011), favorable effects against cancer (Hangen and Bennink, 2002) and decreases diabetes and obesity (Ahn et al., 2013). Furthermore, common beans play a very important role in sustaining soil fertility by fixation of atmospheric nitrogen and organic matter to the soil. It is a dual-purpose crop producing grains as well as fodder for livestock. As a cover crop, it is efficient in suppressing weeds and prevents soil erosion.
Production of common bean in South western (SW) Kenya is constrained by various biotic stresses including insect pests especially pod-borers and weevils which may cause yield loss of up to 80%. There is also lack of cultivars with consumer quality attributes such as taste, palatability and fast cooking. However, the greatest limitation to bean production in SW Kenya are fungal diseases particularly root rots caused by Fusarium, Rhizoctonia and Pythium (Otsyula et al., 2003). Pythium root rot disease can cause yield losses of up to 100% in susceptible varieties. Farmers have abandoned these susceptible cultivars even though they produce high yields. This has led to gradual genetic erosion of once elite landraces. The use of resistant bean varieties is the most effective, economical and environmentally sustainable strategy to control Pythium root rot disease (Binagwa et al., 2016). However, this requires identification of resistant genotypes, and incorporation of the disease resistance into agronomically desirable varieties. There is need to characterize common bean germplasm for Pythium root rot resistance in order to identify markers for selection of resistant varieties. The markers can be used for discarding susceptible germplasm before tolerant varieties are screened in the glasshouse or field and will fast track the development of high yielding Pythium root rot-resistant cultivars for farmers.
Genetic differences that exist between common bean accessions can be associated with economically important traits and used for germplasm characterization. Molecular markers are useful tools in estimation of genetic diversity and identification of alleles of interest without interference from changes in environmental parameters. Gene-based molecular markers represent an important resource for characterization of germplasm and elucidation of gene functions. Peroxidase, a key enzyme in metabolic pathways is an example of gene-based molecular marker in plants. Peroxidases (POXs) belong to a multigene family and exhibit high sequence variability with the existence of conserved domains (Oliva et al., 2009). Conserved DNA regions of peroxidase share the same priming site and are distributed across the genomes of different genotypes in different patterns hence polymorphisms can be detected within species. POXs are glycoprotein enzymes containing heme cofactor and utilize H2O2 in oxidation reactions involving a range of compounds. These enzymes perform diverse roles in plants including detoxification of reactive oxygen species generated during biotic and abiotic stresses (Mittler et al., 2004; Gill and Tuteja, 2010; Zhang et al., 2014), inducing defense response against pathogens (Passardi et al., 2005), formation of lignin and suberin (Herrero et al., 2013), metabolism of auxin, healing of wounds and plant–microbial symbiosis (Passardi et al., 2005). Peroxidases also catalyzes deamination of trans- cinnamic acid in a biosynthesis pathway leading to the formation of phenolic compounds which have many vital activities in plants such as regulation of plant growth differentiation (Vicuna, 2005), inhibition of pathogens (Almagro et al., 2008), and tolerance to abiotic stresses (Gill and Tuteja, 2010; Zhang et al., 2014).
The objective of this study was to characterize common bean landraces from South Western Kenya using peroxidase gene markers for resistance to Pythium root rot disease. This information would be useful in understanding the genetic relationship between different landraces and selection of Pythium root rot-resistant parents for use in present and future breeding programs.
Plant materials
Fifty one common bean landraces were evaluated in the greenhouse for resistance to Pythium root rot disease. These comprised of 25 landraces collected from farmers’ fields in different agro-climatic zones of south western Kenya and 26 germplasm obtained from the National Gene Bank of Kenya (GBK), Muguga, Kenya. The germplasm from GBK were collected also from South Western Kenya region in 1983 - 1984 and preserved.
Preparation and inoculation of Pythium spp. pathogen
Pythium spp. was isolated from symptomatic bean plants collected from farmers’ fields in southwestern Kenya. The bean plants were uprooted washed with running water and stem bases cut off, surface sterilized with 1.5% solution of sodium hypochlorite for 30 s and then rinsed three times with sterile distilled water and blot dried. The cut stem tissues were plated on Potato Dextrose Agar (PDA) media supplemented with 50 ppm streptomycin and incubated for 7 days at 24°C. The Pythium spp. was identified based on morphological and cultural features and confirmed using fungal identification keys as described by Watanabe (2010).
To prepare Pythium inocula, three day old, actively growing hyphal regions measuring 4 mm2 were aseptically cut and grown on autoclaved millet seeds. The culture of Pythium spp. was then mixed with pre-sterilized loam soil in a ratio of 1:8 v/v in wooden flats measuring 48 cm × 72 cm and the inoculum allowed to establish for a period of 14 days in the dark. Two rows of bean seeds (10 seeds per landrace) were planted in the wooden flats and each treatment replicated thrice. The control experiment contained seeds sown in sterilized loam soil without Pythium inoculum. Replications were set using the standard randomized complete block design (RCBD). The experiment was repeated twice.
Pythium root rot disease assessment in the glasshouse
After germination, bean plants were watered after every three days to ensure optimum growth conditions. Thirty six days after planting, seedlings were uprooted and washed with tap water to remove soil from the roots and the plants were individually assessed for disease severity using the CIAT nine-point severity scale (Otsyula et al., 2003). Data on disease severity were subjected to analysis of variance (ANOVA) using the GenStat 11th Edition (VSN International Ltd., 2008) software.
Genomic DNA extraction
Plant genomic DNA was isolated using cetyltrimethyl ammonium bromide (CTAB) method as described by Gyang et al., (2017). Approximately 200 mg of young leaf samples were weighed and crushed in pre-warmed extraction buffer consisting of 700 µl CTAB buffer plus 150 µl 20% sodium dodecyl sulfate (SDS) to form a homogenous paste. The homogenate was transferred into 1.5 ml centrifuge tubes and incubated in a water-bath at 55°C for 20 min. The tubes were gently inverted five times after every five minutes during the incubation period to ensure uniform distribution of the homogenized tissues in the buffer. After incubation, the samples were spun in a micro-centrifuge for 10 min at 13,800 rpm. The debris was discarded and the supernatant transferred to a new 1.5 ml centrifuge tube preceding addition of equal volume of phenol: chloroform: isoamyl alcohol (25:24:1). The contents in the centrifuge tubes were mixed 50 times by inversion to allow proper mixing before spinning for 7 min at 13,800 rpm. The top layer was transferred to new 1.5 ml centrifuge tubes followed by addition of 50 µl of 7.4 M ammonium acetate and 2 volumes of ice cold absolute ethanol. The tubes were incubated at -20°C for 1 h and then centrifuged for 10 min at 10,000 rpm to pellet the precipitated nucleic acids. The supernatant was discarded and 500 µl of a wash solution (75% ethanol and 15 mM ammonium acetate) was added to wash the pellet. The washing step was repeated twice. After every wash, the centrifuge tube was spun for 5 min at 12,000 rpm and the supernatant discarded. The DNA pellet was air dried for 10 min in the fume hood and dissolved in 70 µl TE buffer. Ribonuclease A (3 µl of 10 mg/ml) was added to the dissolved nucleic acids and incubated in a water bath at 37°C for 30 min. The dissolved DNA was stored at -20°C for subsequent molecular analysis.
Peroxidase-gene-based markers and polymerase chain reactions
Primers (Table 1) designed from peroxidase cDNA sequences of Arabidopsis and rice (Welinder et al., 2002; Gulsen et al., 2007; Nemli et al., 2014) were used to detect polymorphism in common bean accessions. Polymerase chain reaction (PCR) reactions were done in a total volume of 20 µl, made up of 5 µl 1× GoTaq Mix (Promega Corporation, Madison, USA), 1 µl of each of the forward and reverse POX markers (10 µM), 1 µl genomic DNA (20 ng), 12 µl nuclease-free water. Amplifications were done in an MJ MiniTM Thermal Cycler machine (Bio-Rad, Singapore) as follows: initial denaturation at 94°C for 5 mins, for 30 s, followed by denaturation (30 s) at 94°C, annealing at 46 - 56°C (45 s), extension (1 min) at 72°C, then a final extension at 4°C for 7 mins. PCR reaction for each POX primer was done at least twice using DNA extracted from different plants of the same landrace and only clear bands that can be reproduced were used during analysis of data.
Gel electrophoresis
The products of the PCR reactions were ran on a 2% agarose gel for 70 mins at 65 Volts. Amplified DNA bands were photographed using a UV transilluminator and scored for further analysis. The fragment sizes were evaluated base on how they moved through the agarose gel in comparison to a 100-bp DNA ladder (Bioneer, South Africa).
Scoring of alleles and data analysis
Each band was scored as present (1) and absent (0) to generate binary matrix for the 5 POX markers. This POX data was used for analysis of genetic diversity and population structure. The polymorphic information content (PIC) was determined for each peroxidase gene locus following the equation PIC = 1 - ∑ (pi)2 which was described by Botstein et al., (1980) (where pi is the population carrying the ith allele). The similarity matrix generated using Nei’s genetic distance (Nei and Li, 1979) was used to construct a dendrogram using the unweighted pair-group method arithmetic mean (UPGMA) by the use of MVSP 3.1 program.
Population genetic structure analysis was done with a clustering approach of a Bayesian model-base, clustering approach in the STRUCTURE version 2.3.4 program (Pritchard et al., 2000). An analysis of all 51 landraces was done using the number of clusters (K) ranging from 1-10, and a burn-in period of 5,000 iterations with 50,000 replications of Markov Chain Monte Carlo (MCMC). Results were not significantly affected with longer burn-in periods. The runs showing the maximum posterior probability for each K value was used. The ad hoc statistic ΔK was used to estimate the total sub-populations, and to determine K (Evanno et al., 2005). Principal component analysis (PCA) was carried out depending on the variation patterns of the POX gene, and a two dimensional representation of relationships across the 51 common bean landraces using XLSTAT program was generated. Analysis of molecular variance (AMOVA) within and among populations was done with the GenAlEx (v6.5) software (Peakall and Smouse, 2012).
Phenotyping of Pythium root rot disease
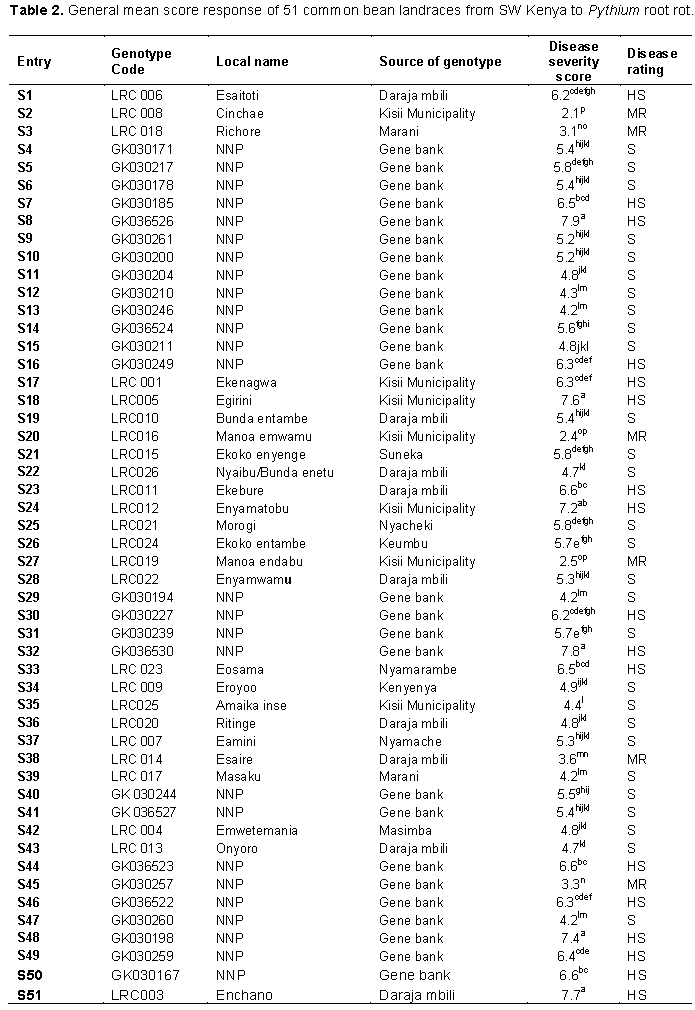
Analysis of variance showed highly significant (p < 0.001) differences in disease severity among the 51 landraces. Average disease severity of all the landraces ranged from 2.1 to 7.8. Based on the disease severity scores against Pythium root rot in the glasshouse, all the 51 common bean landraces were classified in such a way that six (11.77%), twenty-eight (54.90%) and seventeen (33.33%) were moderate-resistant, susceptible and highly susceptible, respectively (Table 2). The landraces with the lowest Pythium root rot disease severity were LRC008, LRC014, LRC016, LRC018, LRC019 and GK030257. None of the 51 landraces was found to be resistant to Pythium root rot disease.
Polymorphism detected using POX markers
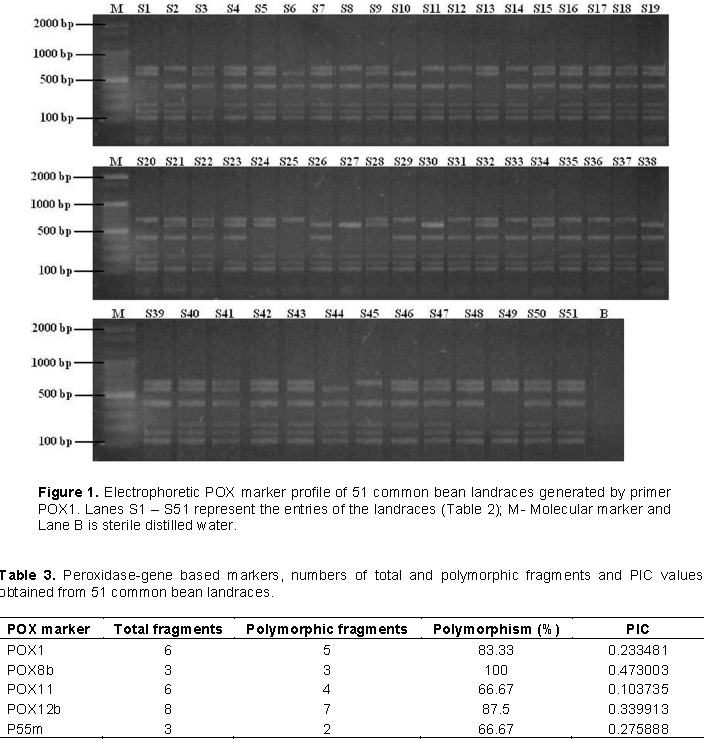
Five POX primers were used to characterize 51 common bean landraces based on the amplification of clear banding patterns. The number of bands generated among the 51 common bean landraces using these 5 primers was 1119, which ranged from 3 (POX8b and POX55m) to 8 (POX12b), with an average of 4.8 bands/primer. A sample amplification pattern of the primer POX1 is shown in Figure 1. Out of the fragments scored, 81% were polymorphic. The percentage polymorphic loci varied from 66.6% (POX 11 and P55m) to 100% (POX 8b) with an average of 80.6% bands /primer (Table 3). Primer POX8b gave 100% polymorphism, indicating the capability of POX primers to detect high levels of polymorphism among common bean (Table 3). The PIC value ranged from 0.10 (POX11) to 0.47 (POX8b).
Genetic relationships among the common bean landraces
A pair-wise comparison among the landraces were used to calculate the genetic similarity coefficient based on the proportion of shared bands. The genetic similarity among the 51 landraces ranged from 0.44 to 1.0, with an average of 0.72. The lowest value of genetic similarity (44%) was observed between S8 (highly susceptible) and S28 (susceptible) common bean landraces. Also relatively low values of genetic similarity was observed between the landraces S4 and S8 (53.8%), S13 and S8 (52%), S24 and S8, (46.2), S27 and S8 (45.8) and; S28 and S8 (45.8%). The maximum genetic similarity (100%) was observed between landraces S7 (highly susceptible) and S19 (susceptible). The genetic similarity matrices showed that the 6 landraces that were moderate resistant were not genetically similar.
Phylogenetic analysis
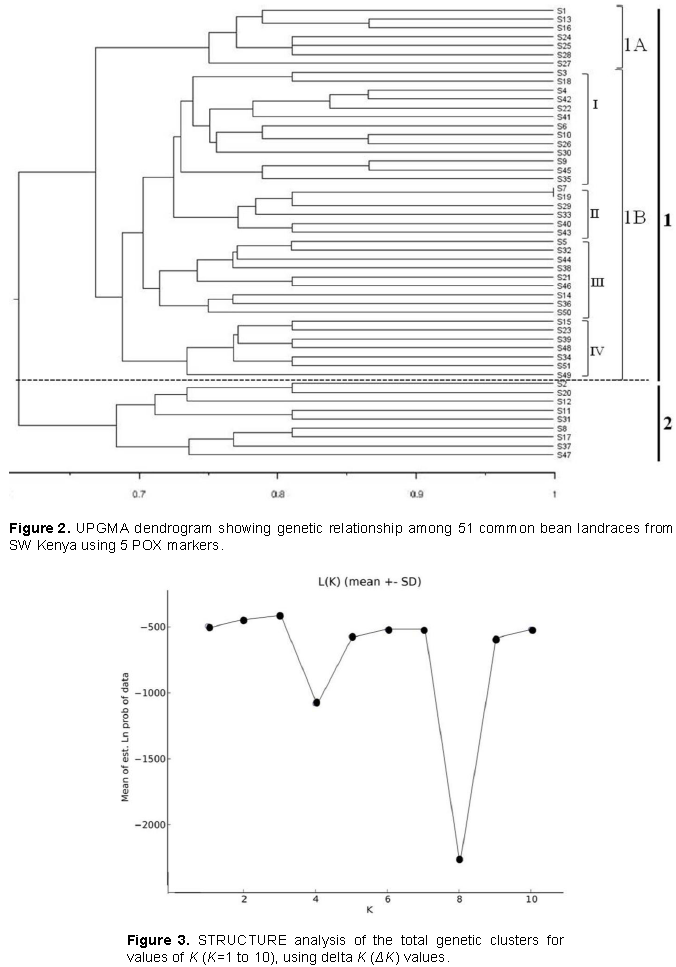
Based on the genetic similarity matrix, a dendrogram was constructed wherein 51 common bean landraces were separated into two major Clusters 1 and 2 (Figure 2). Cluster 1 was the largest and is divided into 2 sub-clusters (namely 1A and B) which in total contained 42 bean landraces of which 4 (S3, S27, S38 and S45) were moderate resistant. Sub-cluster 1Ais the smallest and contains only one (S27) moderate resistant landrace. Sub-cluster 1B was divided into four groups namely I – IV. Groups I and III contained 2 (S3 and S45) and 1 (S38) moderate resistant landraces, respectively, while in Groups II and IV none of the landraces were found to be moderate resistant. Cluster 2 contained 9 landraces of which two (S2 and S20) were moderate resistant. Generally, the landraces form south Western Kenya did not form specific clusters or groups
Population structure analysis
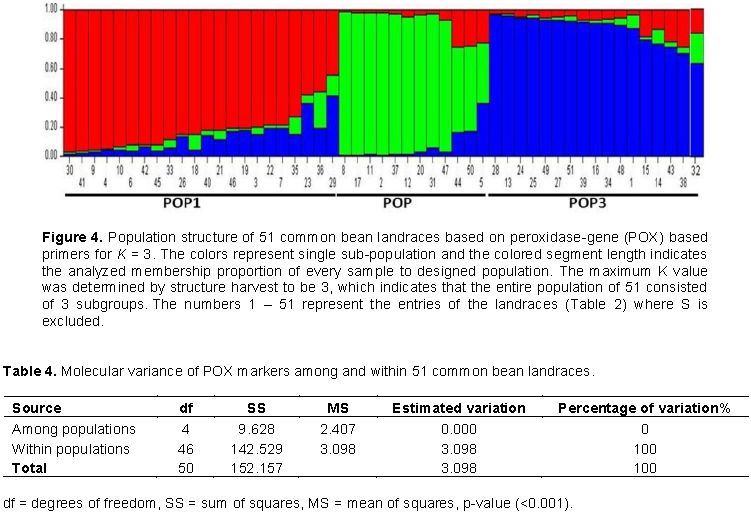
The 51 common bean landraces were analyzed for population structure using Bayesian base method without any prior classification to know the highest populations (K). The peak plateau of adhoc measure ΔK was found to be K = 3 (Figure 3), which indicated that the entire 51 landraces were distributed into three groups (POP1, POP2 and POP3) (Figure 4). The POP2 was the smallest group consisting 12 landraces (23.53%) of which 2, 14 and 6 were found to be moderately resistant, susceptible and highly susceptible, respectively. The POP3 included 17 (33.33%) landraces of which 2, 8 and 7 were found to be resistant, susceptible and highly susceptible, respectively. On the other hand, POP1 was the largest group comprising of 22 (43.14%) landraces of which 2, 6 and 14 were resistant, susceptible and highly susceptible, respectively. There were equal numbers of resistant genotypes (2) in all the three groups. The structure analysis did not differentiate resistant and susceptible landraces into separate groups.
Estimation of population genetics through AMOVA analysis
Analysis of molecular variance was calculated to estimate the partitioning of genetic variance among and within populations. Within population variance explained 100% and no variance (0%) was observed among population (Table 4). All the diversity of common bean landraces from south western Kenya resided within the populations.
The presence of genetic variability in germplasm is a pre-requisite for efficient utilization of available genetic resources for breeding programmes. Understanding genetic differences for Pythium root rot resistance is useful for exploiting landraces as breeding parents. Gene based molecular markers in legumes have been used to identify genotypes, study genetic diversity and determining the phylogenetic relationships (Nemli et al., 2014). This study determined the genetic variations and population structure in common bean landraces from South Western Kenya using peroxidase-gene markers. Plant peroxidase genes serve important roles in providing plants resistance to various biotic stresses resistance (Bela et al., 2015; Mir et al., 2015; Passardi et al., 2005), and peroxidase marker patterns might be utilized in defining relationships among plant genotypes in relation to their adaptive conditions.
In the current study, 51 common bean landraces from South Western Kenya were characterized using five POX loci for resistance to Pythium root rot. The POX markers were previously made using conserved motifs of rice and Arabidopsis peroxidase by Gulsen et al. (2007). The total amplified alleles on each locus varied from 3 - 8, with an average score of 4.8. This compares favorably with Nemli et al. (2014) who reported 1 - 8 alleles on each locus and had an average value of 4.0 when characterizing common bean genotypes from Turkey using POX markers. All the POX markers used in this study were polymorphic. However, the markers had a low average PIC value of 0.28, implying that the landraces used in the present study were closely related. The mean value of PIC observed in this study was lower than that reported by Nemli et al. (2014) and Wittayawannakul et al., (2010), who estimated variation among common bean genotypes and Garcinia species, respectively, using POX polymorphisms and got mean PIC values of 0.40 and 0.79, respectively.
The mean genetic similarity coefficients ranged from 0.44 to 1.0, with an average of 0.72, which indicates high genetic diversity among the 51 landraces. The high genetic diversity in these landraces is a valuable resource for broadening the genetic base in common bean breeding programs. The genetic differences among moderately resistant landraces to Pythium root rot disease as revealed by their clustering into different clusters and groups suggest the presence of different sources and sufficient genetic variation for resistance to Pythium root rot. This genetic variability can be exploited for developing cultivars resistant to Pythium root rot. In the present study, the dendrogram constructed using UPGMA method suggested occurrence of two major clusters and illustrated no clear pattern of distribution of moderate resistant, susceptible and highly susceptible landraces. The relationship between landraces in the cluster groups could not be attributed to their resistance to Pythium root rot. The 51 common bean landraces were divided by STRUCTURE analysis into three groups and did not indicate any distribution pattern in terms of their reaction to Pythium root rot. It is therefore predicted that, combining landraces from the different clusters and groups as parents in breeding programmes would result in broadening Pythium root rot resistance genes in the population.
Incorporating Pythium-moderate resistant landraces which have other desirable agronomic and consumer quality traits such as high iron and zinc content, fast cooking ability, from the different clusters and groups as parents for breeding, would ensure the diversification of resistance to the disease while creating new hybrids. Six common bean landraces from South western Kenya with the potential moderate resistance to Pythium root rot were identified. This study has shown that despite the damaging effects of the disease on the crop, there is a significant amount of genetic variability among the local landraces, which could be utilized in breeding programs targeting development of Pythium root rot-resistant germplasm and cultivars to diversify resistance to the disease. This molecular characterization of common bean landraces from South Western Kenya will give valuable information for breeders and lead to the development of new cultivars for resistance to Pythium root rot. Since the POX markers used in the present study were developed to determine genetic diversity, there is need to use markers linked to the genes responsible for resistance to Pythium root rot in order to facilitate the effective identification of quantitative trait loci linked to this trait.
The authors have not declared any conflict of interests.
The authors thank the National Research Fund (NRF) of Kenya for funding this study. The authors also thank the Department of Biochemistry, University of Nairobi for providing research facilities.
REFERENCES
|
Ahn Y, Park S, Kwack H, Kim MK, Ko K, Kim SS (2013). Rice-eating pattern and the risk of metabolic syndrome especially waist circumference in Korean Genome and Epidemiology Study (KoGES). BMC Public Health 13:61.
Crossref
|
|
|
|
Almagro L, Gomez RLV, Belchi-Navarro S, Bru R, Ros BA, Pedreno MA (2008). Class III peroxidases in plant defense reactions. Journal of Experimental Botany 60(2):377-390.
Crossref
|
|
|
|
|
Bela K, Horváth E, Gallé Á, Szabados L, Tari I, Csiszár J (2015). Plant glutathione peroxidases: emerging role of the antioxidant enzymes in plant development and stress responses. Journal of Plant Physiology 176:192-201.
Crossref
|
|
|
|
|
Binagwa PH, Bonsi CK, Msolla NS, Ritte IP (2016). Morphological and molecular identification of Pythium spp. isolated from common beans (Phaseolus vulgaris) infected with root rot disease. African Journal of Plant Science 10(1):1-9.
Crossref
|
|
|
|
|
Botstein D, White RL, Skolnick M, Davis RW (1980). Construction of a genetic linkage map in man using restriction fragment length polymorphisms. American Journal of Human Genetics 32(3):314-331.
|
|
|
|
|
Evanno G, Regnaut S, Goudet J (2005). Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Molecular Ecology 14(8):2611-2620.
Crossref
|
|
|
|
|
FAOSTAT (2013). Statistical Database, Food and Agriculture.
View
|
|
|
|
|
Gill SS, Tuteja N (2010). Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiology and Biochemistry 48(12):909-30.
Crossref
|
|
|
|
|
Gulsen O, Shearman RC, Heng-Moss TM, Mutlu N, Lee DJ, Sarath G (2007). Peroxidase gene polymorphism in buffalograss and other grasses. Crop science 47(2):767-772.
Crossref
|
|
|
|
|
Gyang PJ, Nyaboga EN, Muge EK (2017). Molecular characterization of common bean (Phaseolus vulgaris L.) genotypes using microsatellite markers. Journal of Advances in Biology and Biotechnology 13(2):1-15.
Crossref
|
|
|
|
|
Hangen L, Bennink MR (2002). Consumption of black beans and navy beans (Phaseolus vulgaris) reduced azoxymethane-induced colon cancer in rats. Nutrition and Cancer 44(1):60-65.
Crossref
|
|
|
|
|
Herrero J, Fernandez-Perez F, Yebra T, Novo-Uzal E, Pomar F, Pedreno MA (2013). Bioinformatic and functional characterization of the basic peroxidase 72 from Arabidopsis thaliana involved in lignin biosynthesis. Planta 237(6):1599-1612.
Crossref
|
|
|
|
|
Mattei J, Hu FB, Campos H (2011). A higher ratio of beans to white rice is associated with lower cardiometabolic risk factors in Costa Rican adults. American Journal of Clinical Nutrition 94(3):869-876.
Crossref
|
|
|
|
|
Mir AA, Park S, Sadat MA, Kim S, Choi J, Jeon J, Lee Y (2015). Systematic characterization of the peroxidase gene family provides new insights into fungal pathogenicity in Magnaporthe oryzae. Scientific Reports 5:11831.
Crossref
|
|
|
|
|
Mittler R, Vanderauwera S, Gollery M, Van Breusegem F (2004). The reactive oxygen gene network in plants. Trends in Plant Science 9(10):490-498.
Crossref
|
|
|
|
|
Nei M, Li WH (1979). Mathematical model for studying genetic variation in terms of restriction endonucleases. Proceedings of the National of Academy of Sciences, USA 76(10):5269-5273.
Crossref
|
|
|
|
|
Nemli S, Kaya HB, Tanyolac B (2014). Genetic assessment of common bean (Phaseolus vulgaris L.) accessions by peroxides gene-based markers. Journal of Science, Food and Agriculture 94(8):1672-1680.
Crossref
|
|
|
|
|
Oliva M, Theiler G, Zamocky M, Koua D, Margis-Pinheiro M, Passardi F, Dunand C (2009). PeroxiBase: a powerful tool to collect and analyse peroxidase sequences from Viridiplantae. Journal of Experimental Botany 60(2):453-459.
Crossref
|
|
|
|
|
Otsyula RM, Buruchara RA, Rubaihayo P (2003). The transmission and inheritance the of root rot fungus (Pythium) resistance to bean genotypes. African Crop Science Society 6:288-297.
|
|
|
|
|
Passardi F, Cosio C, Penel C, Dunand C (2005). Peroxidases have more functions than a Swiss army knife. Plant Cell Reports 24(5):255-265.
Crossref
|
|
|
|
|
Peakall R, Smouse PE (2012). GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research – an update. Bioinformatics 28(19):2537-2539.
Crossref
|
|
|
|
|
Pritchard JK, Stephens M, Donnelly P (2000). Inference of population structure using multilocus genotype data. Genetics 155(2):945-959.
|
|
|
|
|
Vicuna D (2005). The role of peroxidases in the development of plants and their responses to abiotic stresses, Doctoral thesis, Dublin Institute of Technology.
|
|
|
|
|
Watanabe T (2010). Pictorial atlas of soil and seed fungi: morphologies of cultured fungi and key to species. 3rd Edn., CRC Press, Taylor and Francis Group, FL., USA.
Crossref
|
|
|
|
|
Welinder KG, Justesen AF, Kjaersgard IVH, Jensen RB, Rasmussen SK, Jespersen HM, Duroux L (2002). Structural diversity and transcription of class III peroxidases from Arabidopsis thaliana. European Journal of Biochemistry 269(24):6063-6081.
Crossref
|
|
|
|
|
Wittayawannakul W, Garcia RN, Yllano OB, Borromeo TH, Namuco LO, Tecson-Mendoza EM (2010). Assessment of genetic diversity in Garcinia species using peroxidase, RAPD and gene sequence specific amplification polymorphism (GSSAP). Philippines Agricultural Science 93(1):31-41.
|
|
|
|
|
Zhang LX, La JH, Liang ZS, Ashraf M (2014). Interactive effects of sudden and gradual drought stress and foliar-applied glycine-betaine on growth, water relations, osmolyte accumulation and antioxidant defence system in two maize cultivars differing in drought tolerance. Journal of Agronomy and Crop Science 200(6):425-433.
Crossref
|
|