Full Length Research Paper

ABSTRACT
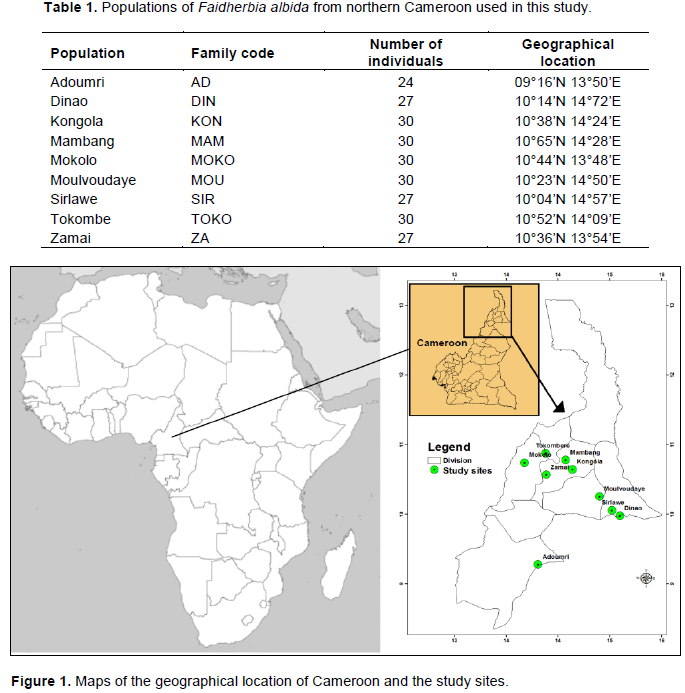
Faidherbia albida (DEL.) A. Chev is widely grown in the northern region of Cameroon because of its many benefits to local communities. However, droughts and deforestation have led to decreases in the abundance of this species, increasing the need to identify improved genotypes for conservation, management, and breeding. The genetic diversity of F. albida in nine populations from the Sudano-Sahelian region of northern Cameroon was characterized using microsatellite (SSRs) markers. A total of 28 alleles were recorded across 8 loci and 255 samples. The effective mean number of alleles per locus was 2.3. Observed heterozygosity ranged from 0.24 to 0.30, while expected heterozygosities ranged from 0.22 to 0.26. For most loci, FIS was negative. Higher variation was observed within than among the northern Cameroon populations, and principal component and admixture analyses did not reveal any population substructure. Phenotypic diversity in 3-month-old seedlings was also characterized and significant within population variation was found for most morphological traits. Although some populations differed significantly for one or more traits, in general the populations were phenotypically similar. These results suggest little barrier to gene flow between populations of F. albida in northern Cameroon, and that no single provenance is likely to be superior for selection and breeding purposes.
Key words: Faidherbia albida, phenotypic, genotypic, variation, populations, breeding.
INTRODUCTION
MATERIALS AND METHODS

RESULTS
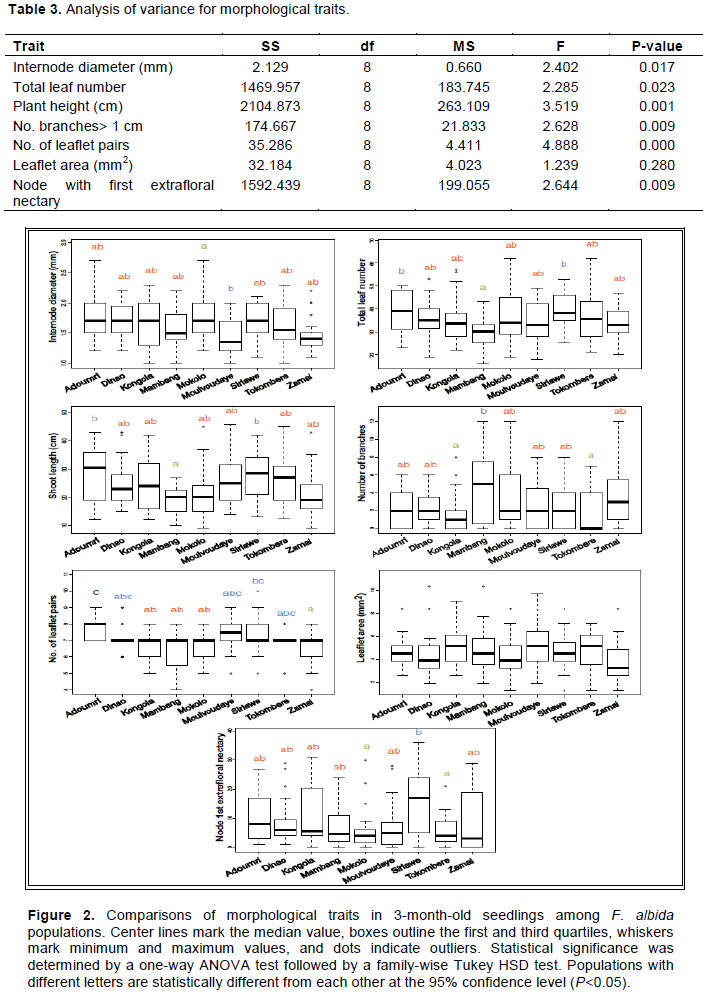

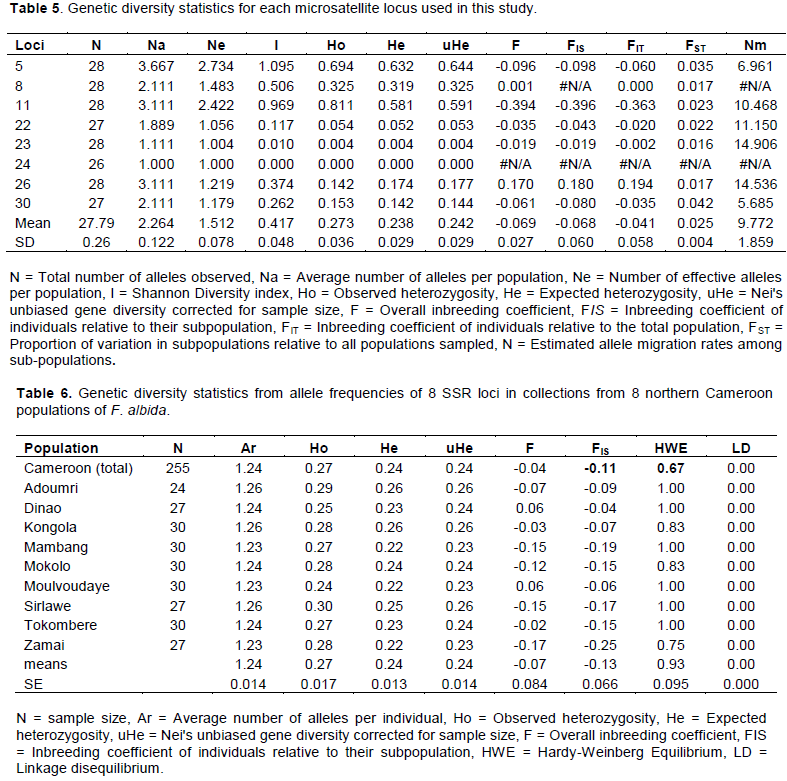
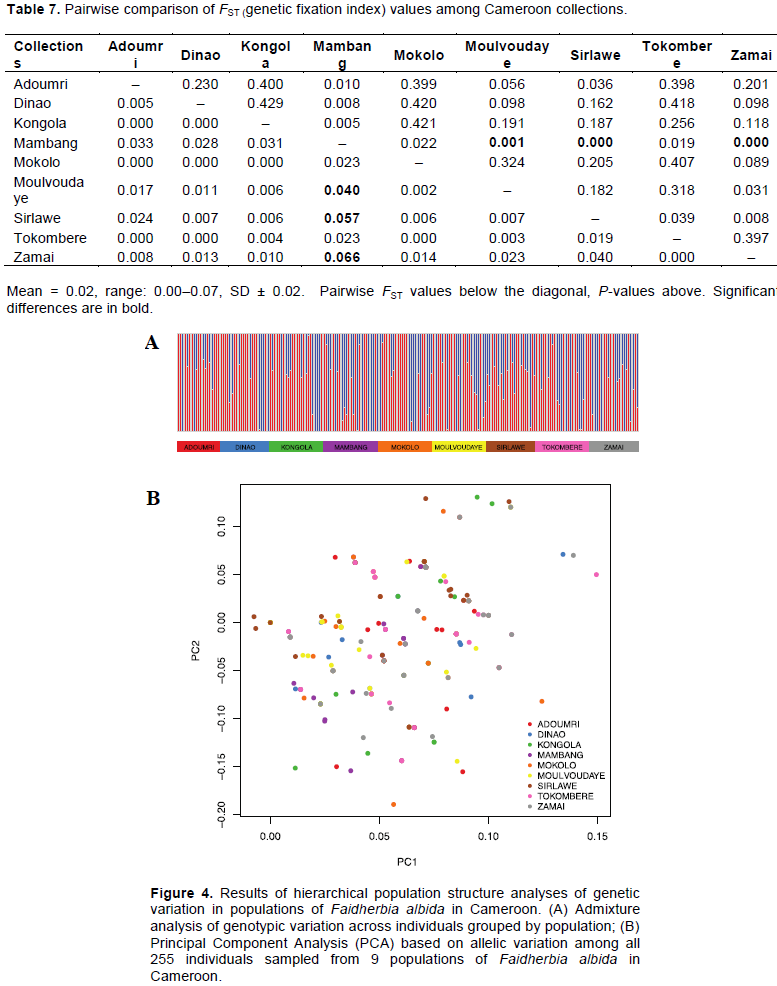
DISCUSSION
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
REFERENCES
|
Abdelazim IM, Fagg CW, Harris SA (1997). Seed and seedling variation amongst provenances in Faidherbia albida. Forest Ecology and Management 97(2):197-205. |
|
|
Alexander DH, Novembre J, Lange K (2009). Fast model-based estimation of ancestry in unrelated individuals. Genome Research 19(9):1655-1664. |
|
|
Alkimim ER, Caixeta ET, Sousa TV, Resende MDV, da Silva FL, Sakiyama NS, Zambolim L (2020). Selective efficiency of genome-wide selection in Coffea canephora breeding. Tree Genetics and Genomes 16(3). |
|
|
Barnes RD, Fagg CW (2003). Faidherbia albida. Monograph and Annotated Bibliography. Tropical Forestry Papers No. 41, Oxford Forestry Institute, Oxford, U.K. |
|
|
Benoit L, Born C, Vignes H, Chevallier MH, Todou G, Debain C, Joly HI (2011). Polymorphic microsatellite loci from Dacryodes edulis (Burseraceae), a Central African rainforest and fruit-tree species. American Journal of Botany 98(4):74-75. |
|
|
Boffa JM (1999). Agroforestry parklands in sub-Saharan Africa (No. 34). Food and Agriculture Organization Conservation Guide 230p. |
|
|
Chang Y, Liu H, Liu M, Liao X, Sahu SK, Fu Y, Song B, Cheng S, Kariba R, Muthemba S, Hendre PS, Mayes S, Ho WK, Yssel AEJ, Kendabie P, Wang S, Li L, Muchugi A, Jamnadass R, Lu H, Peng S, Van Deynze A, Simons A, Yana-Shapiro HYves Van de Peer Y, Xu X, Yang H, Wang J, Liu X (2018). The draft genomes of five agriculturally important African orphan crops. GigaScience 8:1-16. |
|
|
Dangasuk OG, Gudu S, Aman RA, Brandl T (2002). Isozyme variation and growth performance in 16 provenances of African Faidherbia albida (Del.) A. Chev. in Kenya." Discovery and Innovation (special edition) April: 26-36. ISSN 1015-079X |
|
|
Dangasuk GO, Odago PO, Were BA, Guru S (2006). Geographical characterisation of African provenances of Faidherbia albida. African Crop Science Journal 14(4):263-273. |
|
|
Duncan DB (1955). Multiple range and multiple F tests. Biometrics 11(1):1-42. |
|
|
Excoffier L, Smouse PE, Quattro JM (1992). Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data. Genetics 131:479-491. |
|
|
FAO (1974). FAO Panel of Experts in Forest Gene Resources Report of the Third Session. Rome, Italy 90p. |
|
|
Franzel S, Jaenicke H, Janssen W, Ayuk E, Boland D (1996). Choosing the right trees: Setting priorities for multipurpose tree improvement. ISNAR Research Report 8:87p. |
|
|
Garcia F, Noyer JL, Risterucci AM, Chevallier MH (2004). Genotyping of Mature Trees of Entandrophragma cylindricum with Microsatellites. Journal of Heredity 95(5):454-457. |
|
|
Gautier D, Mana J, Rocquencourt A, Tapsou, Njiti C (2002). Faut-il poursuivre l'opération Faidherbia du DPGT au Nord Cameroun. Savanes africaine: des espaces en mutation, des acteurs face à de nouveaux défis. Actes du colloque, mai 2002, Prasac, Maroua, Cameroun, N'Djamena, Tchad 9 p. ⟨hal-00133790⟩ |
|
|
Grattapaglia D (2017). Status and perspectives of genomic selection in forest tree breeding. In Genomic selection for crop improvement. Springer, Cham pp. 199-249. |
|
|
Guries RP, Ledig TF (1981). Genetic Structure of Populations and Differentiation in Forest Trees. Symposium on Isozymes of North American Forest Trees and Forest Insects, July 27, 1979, Berkeley, California. Pacific Southwest Forest and Range Experiment Station (Berkeley, Calif.). General Technical Report PSW-48:42-47. |
|
|
Harris SA, Fagg CW, Barnes RA (1997). Isozyme variation in Faidherbia albida (Leguminosae Mimosoideae). Plant Systematic and Evolution 207:119-132. |
|
|
Hauser TP (1994). Germination, predation and dispersal of Acacia albida seeds. Oikos 71:421-426. |
|
|
Hedrick PW, Kalinowski ST (2000). Inbreeding depression in conservation biology. Annual Review of Ecology and Systematics 31:139-162. |
|
|
Joly HI, Zeh-Nlo M, Danthu P, Aygalent C (1992). Population genetics of an African Acacia: Acacia albida. Genetic diversity of populations from West Africa. Australian Journal of Botany 40:59-73. |
|
|
Kumar S, Tamura K, Nei M (1994). MEGA: Molecular evolutionary genetics analysis software for microcomputers. Bioinformatics 10(2):189-191. |
|
|
Muona O (1990). Population genetics in forest tree improvement. Plant population genetics, breeding, and genetic resources pp. 282-298. |
|
|
Muchugi A, Kadu K, Kindt R, Kipruto H, Lemurt S, Olale K, Nyadoi P, Dawson I, Jamnadass R (2008). Molecular Markers for Tropical Trees. ICRAF Technical Manual 9:91p. |
|
|
Omondi SF, Kireger E, Dangasuk OG, Chikamai B, Odee DW, Cavers S, Khasa DP (2010) Genetic Diversity and Population Structure of Acacia senegal (L) Willd. in Kenya. Tropical Plant Biology 3(1):59-70. |
|
|
Peakall R, Smouse PE (2012). GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 28(19):2537-2539. |
|
|
Peltier R, Eyog-Matig O (1988). Les essais d'agroforesterie au Nord-Cameroun. Bois et Forets des Tropiques 217(3):3-31. |
|
|
Peltier R (1996). Les parcs à Faidherbia. Cahier scientifique n12, CIRAD, Montpellier, France 312p. |
|
|
Ribaut JM, De Vicente MC, Delannay X (2010). Molecular breeding in developing countries: Challenges and perspectives. Current Opinion in Plant Biology 13(2):213-218. |
|
|
Roupsard O, Ferhi A, Granier A, Pallo F, Depommier D, Mallet B, Joly HI, Dreyer E (1999). Reverse phenology and dry-season water uptake by Faidherbia albida (Del.) A. Chev. in an agroforestry parkland of Sudanese West Africa. Functional Ecology 13(4):460-472. |
|
|
Russell JP, Hedley PE, Dancey LCS, Morris J, Booth A, Odee D, Mwaura L, Omondi W, Angaine P, Machua J, Muchugi A, Iain Milne, Kindt R, Jamnadass R, Dawson Ian K (2014). TropiTree: An NGS-Based EST-SSR Resource for 24 Tropical Tree Species. PLoS ONE 9(7):e102502. |
|
|
Sniezko RA, Stewart HTL (1989). Range-wide provenance variation in growth and nutrition of Acacia albida seedlings propagated in Zimbabwe. Forest Ecology and Management 27:179-197. |
|
|
Tchatchoua DT, Kolyang MG, Caspa RG, Basga SD, Youri A (2019). Variation in seed and seedling traits of Faidherbia albida populations in the Sudano-Sahelian zone of Cameroon. International Journal for Biological and Chemical Sciences 13(4):2029-2040. |
|
|
Tchoundjeu Z, Weber J, Guarino L (1998). Germplasm collections of endangered agroforestry tree species: The case of Prosopis africana in the semi-arid lowlands of West Africa. Agroforestry Systems 39:91-100. |
|
|
van Nocker S, Gardiner SE (2014). Breeding better cultivars, faster: Applications of new technologies for the rapid deployment of superior horticultural tree crops. Horticulture Research 1(1):1-8. |
|
|
Vandenbeldt RJ (1991). Rooting systems of western and southern African Faidherbia albida (Del.) A. Chev. (syn. Acacia albida Del.)--A comparative analysis with biogeographic implications. Agroforestry Systems 14:233-244. |
|
|
Vandenbeldt RJ (1992). Faidherbia albida in the West African semi-arid tropics: Proceedings of a Workshop, 22-26 Apr 1991, Niamey, Niger. World Agroforestry Centre 212 pp. ISBN: 92-9066-220-4 |
|
|
Varshney RK, Graner A, Sorrells ME (2005). Genic microsatellite markers in plants: Features and applications. TRENDS in Biotechnology 23(1):48-55. |
|
|
Watson D (2019). Adaption to Climate Change: Climate Adaptive Breeding of Maize, Wheat and Rice. In Sustainable Solutions for Food Security. Springer, Cham pp. 67-89. |
|
|
Wickens GE (1969). A study of Acacia albida DEL. (Mimosoideae). Kew Bull 23:181-202. |
|
|
World Agroforestry Centre (2009). PRESS RELEASE: Unique Acacia Tree Could Nourish Soils and Life in Africa. |
|
|
Wright JW (1962). Genetics of forest tree improvement. Genetics of forest tree improvement 399 pp. |
|
|
Zargar SM, Raatz B, Sonah H, Bhat JA, Dar ZA, Agrawal GK, Rakwal R (2015). Recent advances in molecular marker techniques: Insight into QTL mapping, GWAS and genomic selection in plants. Journal of crop science and biotechnology 18(5):293-308. |
|
|
Zobel BJ, Talbert JT (1984). Applied forest tree improvement. Wiley and Sons, Inc., New York 505p. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


