Full Length Research Paper

ABSTRACT
Several thrips attractants were screened for compatibility with Metarhizium anisopliae (Metchnikoff) Sorokin (Hypocreales: Clavicipitaceae) and a subset of these for attraction to Megalurothrips sjostedti Trybom (Thysanoptera: Thripidae). Conidial germination and germ tube length of M. anisopliae were used as indicators of its compatibility with thrips attractant. Conidial germination and germ tube length differed significantly according to volatiles of different attractants. The highest conidial germination (76.5±3.5%) and longest germ tube length (130.3±13.4 µm) were recorded in the control, followed by methyl anthranilate (63.8±3.8%; 103.8±8.4 µm), cis-jasmone (61.8±5.9%; 93.8±14.4 µm) and trans-caryophyllene (57.7±6.5%; 96.3±15.5 µm) which were found compatible with M. anisopliae. A Pearson correlation test indicated a significant positive correlation between conidial germination and germ tube length (r =0.6; P<0.0001). The attraction of M. sjostedti to selected thrips attractant also varied significantly among the attractants. Under field conditions, methyl anthranilate was equally attractive to M. sjostedi as Lurem-TR and could be recommended as a thrips attractant that can be combined with M. anisopliae in autoinoculation devices for potential control of M. sjostedti.
Key words: Semiochemicals, conidial germination, germ tube length, Megalurothrips sjostedti, attraction, persistence, field.
INTRODUCTION
In many flower dwelling thrips, host finding is linked to visual, odour and morphological (shape) cues (Rieske and Raffa, 2003; Mainali and Lim, 2011). Subsequently, semiochemical-based products such as Lurem-TR and Thripline have been developed for use in thrips monitoring and management (Sampson and Kirk, 2013; Teulon et al., 2014; Broughton et al., 2015). These semiochemicals can be integrated with other control strategies to improve thrips management in horticulture (Suckling et al., 2012; Sampson and Kirk, 2013).
Entomopathogenic fungi (EPF) are among the alternatives to synthetic chemical pesticides being considered for the management of thrips in horticulture (Ekesi and Maniania, 2007). EPF are generally applied through inundative spray, which requires high amount of inocula, thereby enhancing its cost (Jaronski, 2010). Further, the persistence of conidia applied on foliage is challenged by several environmental parameters such as UV light, rain, temperature (Inglis et al., 2000; Jaronski, 2010). The use of “lure and kill” strategy using autoinoculation device or spot spray application could reduce the amount of inoculum, the cost and sustain fungal persistence in the field (Dimbi et al., 2003; Nana et al., 2014; Mfuti et al., 2016). However, the success of this technology depends on the use of powerful attractants and their compatibility with the entomopathogens. For example, the tick attraction-aggregation-attachment pheromone (AAAP) could attract adult ticks from a distance of 6 m (Nchu et al., 2009) but could not be used in combination with conidia of Metarhizium anisopliae (Metschnikoff) Sorokin (Hypocreales: Clavicipitaceae) because of inhibition of fungal conidia by the pheromone (Nana et al., 2012). Niassy et al. (2012a) and Mfuti et al. (2016) have also reported inhibitory effects of conidia of M. anisopliae by the semiochemical Lurem-TR in autoinoculation device in a screenhouse and field experiments.
Considering the growing interest in integrating attractants with EPF in thrips management (Niassy et al., 2012a; Mfuti et al., 2016), there is a need to identify compounds that are both attractive to thrips and compatible with EPF. The objective of the present study was therefore to identify thrips attractants that are compatible with M. anisopliae in terms of conidial germination and germ tube length since the latter plays a crucial role in fungal infection (Ortiz-Ribbing and Williams, 2006).
MATERIALS AND METHODS
Thrips attractants
Seven compounds used for thrips attraction or with potential attraction for thrips were tested for their compatibility with M. anisopliae isolate ICIPE 69. They were selected on the basis of structural analogies to known attractant such as methyl isonicotinate (Lurem-TR) (Teulon et al., 2007, 2010) but also based on previous studies of Koschier et al. (2000). Information on their chemical characteristics and manufacturers is presented in Table 1. The commercial attractant, Lurem-TR which was earlier reported to be toxic with the entomopathogen (Niassy et al., 2012a) was included in the study as a reference. It is a commercial product which quantity and release rate is standardized; therefore, could not be diluted. In preliminary bioassays, no significant effect of different concentrations (0.1, 10 and 100%) of attractants was observed on conidial germination and subsequently only the recommended concentration of 10% of the pure product was used in the screening bioassays. The pure concentration of all attractants was diluted in paraffin oil.
Crop
Cowpea, Vigna unguiculata L. Walp variety Ken-Kunde1, was planted in 80 m2 plots with an inter- and intra- row spacing of 10 and 45 cm, respectively, in Mbita Thomas Odhiambo Campus (ITOC) (0° 26’ 06.19” S, 34° 12’ 53.13” E; 1,137 above sea level) earlier during rainy season (March 2014). The size of the cowpea farm was about 94 × 22 m. The field experiment of selected attractants was conducted during flowering stage of the crop (45 days after planting). No fertilizers, organic matter or synthetic chemical insecticides were applied during the experiment.
Fungal culture
M. anisopliae isolate ICIPE 69 was obtained from the Arthropod Germplasm Centre of icipe. It is currently commercialized as Campaign® by the Real IPM Ltd, Kenya, for the control of thrips, papaw mealy bug and fruit flies (http://www.realipm.com). It was cultured on Sabouraud Dextrose Agar (SDA) in 9 cm Petri dishes and incubated at 25 ± 2°C in complete darkness. Conidia were harvested from three week-old culture by scraping the surface using a spatula. Conidia were suspended in 10 ml sterile distilled water containing 0.05% Triton X-100 in universal bottles containing glass beads. Conidial suspensions were vortexed for 5 min to produce a homogeneous suspension. Spore concentrations were determined using a haemocytometer.
Effect of thrips attractants on conidial viability and germ tube length of M. anisopliae
The conidial suspension was prepared as described earlier and titrated to 1×107 conidia ml-1. The spores were retained on a nitrocellulose filter membrane (diameter 47 mm, pore size 0.45 µm, Sigma Chemicals) by pouring 10 ml suspension through a filter holder unit (MFS) under aspirator vacuum (Maniania, 1994). The nitrocellulose filter membranes were dried for 30 min under a laminar flow cabinet and transferred to glass desiccators (2.5 L) for exposure to the attractant volatile. Cotton wicks were soaked in 0.5 ml suspensions of each attractant diluted in paraffin oil and placed in desiccators to allow volatile diffusion. Cotton wicks were used as dispenser (Sidahmed et al., 2014). Fungus-treated nitrocellulose membranes were exposed to different thrips attractants and sampled for viability observation at different time intervals of 1, 2, 3, 6 and 8 days. An untreated control without thrips attractant was included. The commercial thrips attractant, Lurem-TR was included as a check. Treatments were randomized and the experiment repeated three times over time.
To determine conidial germination, nitrocellulose filter membranes containing conidia were removed from the desiccators and transferred into 10 ml sterile distilled water containing 0.05% Triton X-100 and vortexed for 3 min to dislodge conidia. Suspension (0.1 ml) titrated to 3 × 106 conidia ml-1 was spread-plated on SDA plates. Plates were incubated at 26 ± 2°C, L12: D12 photoperiod and examined after 18 to 24 h for conidial germination and germ tube length thereafter. Samples that could not be processed the same day were fixed by pouring a drop of lactophenol cotton blue onto the plate to stop further growth. Percentage germination was determined by counting approx. 100 spores per plate under a microscope Leica DMLB at 40 X magnification. The length of germ tubes was measured using a Leica Application Suite (LAS EZ V1.5.0). Average germ tube lengths were obtained from 5 spores taken at random in each cover slip (22 x 22 mm) and replicated three times.
Effect of selected thrips attractants on the attraction of M. sjostedti
Attractants that were found compatible with M. anisopliae from the screening experiment were selected for field experiment to evaluate the attraction of Bean Flower Thrips (BFT), M. sjostedti (Trybom) (Thysanoptera: Thripidae) on cowpea. Lurem-TR was included as reference. Attractants were diluted in Paraffin oil as indicated above. Each attractant suspension was poured in 5 ml Eppendorf tube and suspended in the middle surface of the blue sticky card (10 × 25 cm) (Plate 1). The two items were placed at 30 cm above ground level. Blue sticky cards were separated 10 m from one another to avoid interference. An untreated blue sticky card with no attractant was used as a control. The experiment was conducted during flowering and podding stages of cowpea when BFT populations are high (Ezueh, 1981; Nyasani et al., 2013). Cards were replaced every three days. Numbers of adult BFT were recorded on each card and the experiment was replicated four times over time.

Statistical analysis
Data on conidial germination of M. anisopliae were normalized by arcsine transformation before subjecting them to linear mixed model. Data on M. anisopliae conidial germ tube length and M. sjostedti catches were also analyzed using linear mixed model. Means were separated using Student–Newman–Keuls (SNK) test. A Pearson correlation analysis was carried out to relate conidial viability with the germ tube length. All data analyses were performed using R (R Development Core Team, 2014). The level of significance was maintained at 95%.
RESULTS
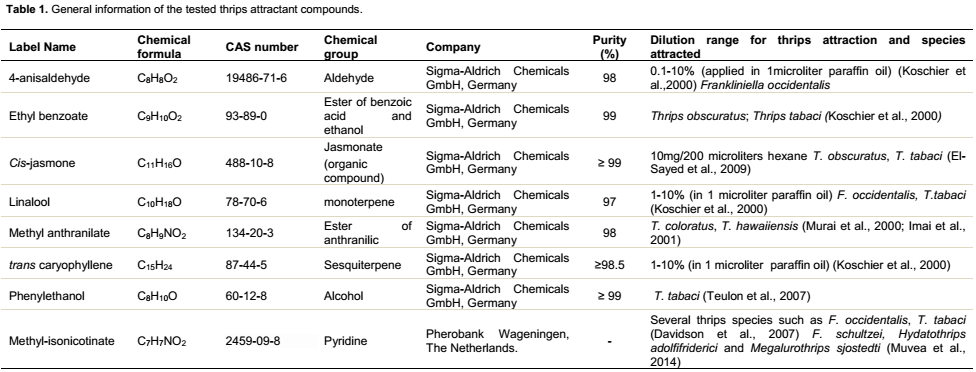
Effect of thrips attractants on conidial viability and germ tube length of M. anisopliae
Overall, the effects of thrips attractants on germination of conidia of M. anisopliae varied significantly between the attractants (F9,268 = 22.1; P<0.0001) (Table 2). The interaction day × attractant was statistically significant (F9,268 = 3.8; P<0.0001). The time of exposure had significant effects on conidial germination, except at 1 day post-exposure when no significant effect was observed (F9,45 = 1.5; P = 0.2) (Table 3). Significant reduction in conidial germination was observed in all the treatments from day 2 (F9,45 = 6.1; P<0.0001), day 3 (F9,45 = 6.8; P<0.0001), Day 6 (F9,45 = 8.3; P<0.0001) and day 8 post-exposure (F9,45 = 8.7; P<0.0001) (Table 3). The conidial germination was significantly higher in the control (62.5 ± 10.0%), followed by cis-jasmone (44.8 ± 16.6%), Solvent (paraffin oil) (42.8 ± 11.0%), methyl anthranilate (36.6 ± 8.0%) and trans-caryophyllene (31.3 ± 16.8%) treatments after 8 days of exposure and was significantly different (Table 3). No conidial germination was observed in Lurem-TR treatment at day 8 post-exposure (Table 3).

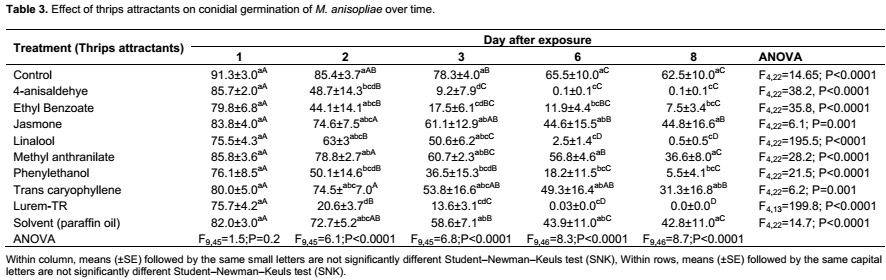
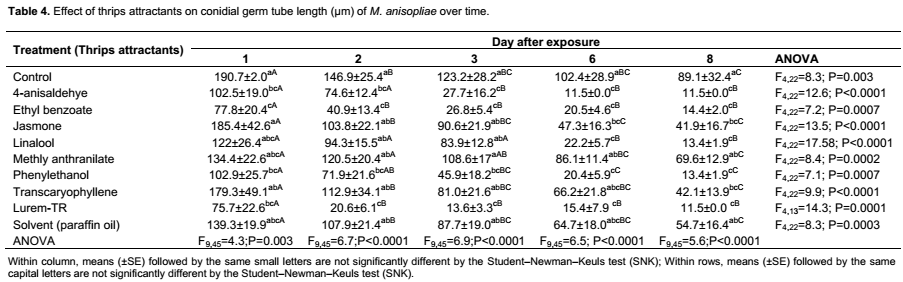
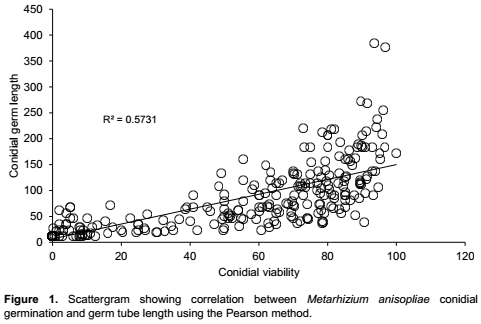
The effect of thrips attractants on germ tube length followed the same trend as with conidial germination where treatments differed significantly (F9,268 = 12.6; P<0.0001) (Table 2). The interaction day × attractant was not statistically significant (F9,268 = 1.0; P=0.5). Exposure time had significant effects on length of the germ tube of M. anisopliae at day 1 (F9,45 = 4.3; P = 0.003), day 2 (F9,45 = 6.7; P<0.0001), day 3 (F9,45 = 6.9; P<0.0001 ), day 6 (F9,45 = 6.5; P<0.0001) and day 8 (F9,45 = 5.6; P<0.0001) post-exposure (Table 4). The highest germ tube length was observed in the control treatment (89.1±32.4 µm) followed by methyl anthranilate (69.6±12.9 µm), solvent (paraffin oil) (54.7 ± 16.4 µm), trans-caryophyllene (42.1 ± 13.9 µm) and cis-jasmone (41.9 ± 16.7 µm) and was not significantly different at day 8 post-exposure (Table 4). No germ tube developed in Lurem-TR treatment at day 6 and 8 post-exposure (Table 4). A significant correlation was found between conidial germination and germ tube length of M. anisopliae (r = 0.6; P<0.0001) (Figure 1).
Effect of selected thrips attractants on the attraction of M. sjostedti
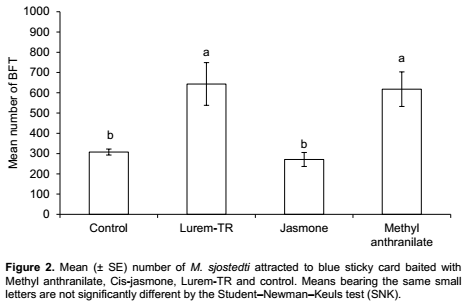
The number of adult BFT caught on the baited sticky cards varied significantly (F3,28 = 7.9; P< 0.0001) between the treatments. More adult BFT were caught on sticky cards treated with Lurem-TR and methyl anthranilate than cards treated with cis-jasmone (Figure 2). No significant difference in thrips catches was found between Lurem-TR and methyl anthranilate. Similarly, there was no significant difference between cis-jasmone and control treatments (Figure 2).
DISCUSSION
Most studies on compatibility of M. anisopliae have focused on agrochemicals and botanicals (Nana et al., 2012; Niassy et al., 2012b) overlooking the potential of semiochemical attractants in insect pest management (IPM). However, a study on compatibility between attractants and EPF is required before their integration in an IPM strategy.
Thrips attractants tested in the present study affected conidial germination and germ tube length differently, with time of exposure being the determining factor. Three out of eight attractants tested namely methyl anthranilate, cis-jasmone and trans-caryophyllene, did not have deleterious effects on conidial germination and germ tube length of M. anisopliae at day 8 post-exposure (Tables 3 and 4). Interestingly, these three attractants have been reported elsewhere to have effects on fungal pathogens. For instance, methyl anthranilate has been reported to significantly reduce the growth of strawberry pathogens such as Botrytis cinerea (Helotiales: Scerotiniaceae), Colletotrichum gloeosporioides and C. acutatum (Glomerellale: Glomerellaceae). In addition, medium supplemented with methyl anthranilate resulted in complete cessation of growth in those pathogens (Chambers et al., 2013).
Recent evidence suggests that jasmonic acid is involved in the induction of genes that act primarily in defense against plant pathogens rather than insects (Halim et al., 2006). Jasmonic acid is part of the plant’s alarm system and defense mechanism. It is a volatile (gas phase of Cis-jasmone) which is released during insect attack and controls the response to damage (Menzel et al., 2014).
Essential oil from Perovskia atriplicifolia Benth (Lamiales: Liamiaceae) containing 9.30% of trans-caryophyllene are reported to have antimicrobial activity against fungal strains (Erdemgil et al., 2007). The difference between these results and our results could be explained by the fact that attractants were used as volatiles in our study while they were used as oil supplements in culture media.
This study also confirmed previous findings on the antifungal effect of Lurem-TR on conidial germination (Niassy et al., 2012a). More recently, it was demonstrated that direct exposure of fungus without separation from Lurem-TR recorded the lowest conidial germination as compared with the other treatments where separation was made. However, fungal persistence increased with distance of separation of Lurem-TR (Mfuti et al., 2016).
The strong correlation observed between conidial germination and germ tube length suggests that fungal inoculum would still cause infection in the insects. The role of germ tube formation in the pathogenesis is well established (Ortiz-Ribbing and Williams, 2006). For instance, comparing four different growth stages of Isaria fumosorosea (Paecilomyces fumosoroseus) (Eurotiales: Trichocomaceae) (conidia, germinated conidia with either one or two germ tubes and hyphal bodies), Fargues et al. (1994) found that germinated conidia and hyphal bodies were more aggressive than ungerminated conidia against first-instar larvae of Spodoptera frugiperda Smith (Lepidoptera: Noctuidae).
The catches of BFT were significantly higher on blue sticky cards baited with methyl anthranilate and Lurem-TR than the control and cis-jasmone baited cards. The increased attraction of BFT to Lurem-TR and blue sticky traps was reported by (Muvea et al., 2014). No difference in BFT attraction was found between the two compounds. Methyl anthrinalate has been reported to be attractive to four species of flower thrips, Thrips hawaiiensis, Thrips coloratus, Thrips flavus, and Megalurothrips distalis, irrespective of sex (Murai et al., 2000; Imai et al., 2001). However, this study is the first report on BFT response to methtyl anthranilate. This study has identified methyl anthranilate as an attractant effective for BFT and also compatible with conidia of M. anisopliae and hence can be considered for a “lure and kill” management strategy for BFT. The “lure and kill” strategy could be adopted either as an autoinoculation device or spot spray. Further studies need to be carried out to validate this proof of concept.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
This study was funded by the African Union through the African Union Research Grant Contract no: AURG/108/2012 and the BMZ (The German Federal Ministry for Economic Cooperation and Development) through GIZ (Deutsche Gesellschaft fur Internationale Zusammenarbeit) Project number: 11.7860.7-001.00, Contract number: 81141840. We acknowledge icipe, Duduville, Nairobi, Kenya and icipe-ITOC, Mbita, Kenya for field facilities on the campuses. We are grateful to Mrs. Barbara Obonyo, Messrs Pascal Oreng, Daniel Ouma, Eleisha Orima and Gregory Chebire for technical assistance. We are grateful to the German Academic Exchange Services (DAAD) through the African Regional Postgraduate Program in Insect Science (ARPPIS) of ICIPE and the African Union Project on Grain Legumes for financial support of the study.
REFERENCES
|
Broughton S, Cousins DA, Rahman T (2015). Evaluation of semiochemicals for their potential application in mass trapping of Frankliniella occidentalis (Pergande) in roses. Crop Prot. 67(1):130-135. |
|
|
Chambers AH, Evans SA, Folta KM (2013). Methyl anthranilate and γ-decalactone inhibit strawberry pathogen growth and achene Germination. J. Agric. Food Chem. 61(51):12625-12633. |
|
|
Davidson MM, Butler RC, Winkler S, Teulon DAJ (2007). Pyridine compounds increase trap capture of Frankliniella occidentalis (Pergande) in a covered crop. New Zealand Plant Prot. 60:56-60. |
|
|
Dimbi S, Maniania NK, Lux SA, Ekesi S, Mueke JM (2003). Pathogenicity of Metarhizium anisopliae (Metsch.) Sorokin and Beauveria bassiana (Balsamo) Vuillemin to three adult fruit fly species: Ceratitis capitata (Weidemann), C. rosa var. fasciventris Karsch and C. cosyra (Walker) (Diptera: Tephritidae). Mycopathology 156(4):375-382. |
|
|
Ekesi S, Maniania NK (2007). Use of entomopathogenic fungi in biological pest management published by Research Signpost, Kerala, India. 333 p. |
|
|
El-Sayed AM, Mitchell VJ, McLaren GF, Manning LM, Bunn B, Suckling DM (2009). Attraction of New Zealand flower thrips, Thrips obscuratus, to cis-jasmone, a volatile identified from Japanese honey suckle flowers. J. Chem. Ecol. 35(6):656-663. |
|
|
Erdemgil FZ, Ilhan S, Korkmaz F, Kaplan C, Mercangoz A, Arfan M, Ahmad S (2007). Chemical composition and biological activity of the essential oil of Perovskia atriplicifolia from Pakistan. Pharm. Biol. 45(4):324-331. |
|
|
Ezueh MI (1981). Nature and significance of pre-flowering damage by thrips to cowpea. Entomol. Exp. Appl. 29(3):305-312. |
|
|
Fargues J, Maniania NK, Delmas JC (1994). Infectivity of propagules of Paecilomyces fumosoroseus during in vitro development to spodoptera frugiperda. J. Invertebr. Pathol. 64(3):173-178. |
|
|
Halim VA, Vess A, Scheel D, Rosahl S (2006). The role of salicylic acid and jasmonic acid in pathogen defence. Plant Biol. 8(3):307-313. |
|
|
Imai T, Maekawa M, Murai T (2001). Attractiveness of methyl anthranilate and its related compounds to the flower thrips, Thrips hawaiiensis (Morgan), T. coloratus Schmutz, T. flavus Schrank and Megalurothrips distalis (Karny) (Thysanoptera: Thripidae). Appl. Entomol. Zool. 36(4):475-478. |
|
|
Inglis GD, Ivie TJ, Duke GM, Goettel MS (2000). Influence of rain and conidial formulation on persistence of Beauveria bassiana on potato leaves and Colorado potato beetle larvae. Biol. Control 18(1):55-64. |
|
|
Jaronski ST (2010). Ecological factors in the inundative use of fungal entomopathogens. Biocontrol 55(1):159-185. |
|
|
Koschier EH, de Kogel WJ, Visser JH (2000). Assessing the attractiveness of volatile plant compounds to Western flower thrips Frankliniella occidentalis. J. Chem. Ecol. 26(12):2646-2655. |
|
|
Mainali BP, Lim UT (2011). Behavioral response of western flower thrips to visual and olfactory cues. J. Insect Behav. 24(6):436-446. |
|
|
Maniania NK (1994). A laboratory technique for infecting adult Tsetse with a fungal pathogen. Insect Sci. Appl. 15(4/5):421-426. |
|
|
Menzel TR, Weldegergis BT, Anja D, Boland W, Gols R, Van Loon JJA, Dicke M (2014) Synergism in the effect of prior jasmonic acid application on herbivore-induced volatile emission by Lima bean plants: transcription of a monoterpene synthase gene and volatile emission. J. Exp. Bot. 65(17):4821-4831. |
|
|
Mfuti DK, Subramanian S, Van Tol RWHM, Wiegers GL, de Kogel WJ, Niassy S, Du Plessis H, Ekesi S, Maniania NK (2016). Spatial separation of semiochemical Lurem-TR and entomopathogenic fungi to enhance their compatibility and infectivity in an autoinoculation system for thrips management. Pest Manage. Sci.72(1):131-139. |
|
|
Murai T, Imai T, Maekawa M (2000). Methyl anthranilate as an attractant for two thrips species and the thrips parasitoid Ceranisus menes. J. Chem. Ecol. 26(11):2557-2565. |
|
|
Muvea AM, Waiganjo MM, Kutima HL, Osiemo Z, Nyasani JO, Subramanian S (2014). Attraction of pest thrips (Thysanoptera: Thripidae) infesting French beans to coloured sticky traps with Lurem-TR and its utility for monitoring thrips populations. Int. J. Trop. Insect Sci. 34(3):197-206. |
|
|
Nana P, Maniania NK, Maranga RO, Boga HI, Kutima HL, Eloff JN (2012). Compatibility between Calpurnia aurea leaf extract, attraction aggregation, and attachment pheromone and entomopathogenic fungus Metarhizium anisopliae on viability, growth, and virulence of the pathogen. J. Pest Sci. 85(2012):109-115. |
|
|
Nana P, Nchu F, Ekesi S, Boga HI, Kamtchouing P, Maniania NK (2014). Efficacy of spot-spray application of Metarhizium anisopliae formulated in emulsifiable extract of Calpurnia aurea in attracting and infecting adult Rhipicephalus appendiculatus ticks in semifield experiments. J. Pest Sci. 88(3):613-619. |
|
|
Nchu F, Maniania NK, Toure´ A, Hassanali A, Eloff JN (2009). The use of a semiochemical bait to enhance exposure of Amblyomma variegatum (Acari: Ixodidae) to Metarhizium anisopliae (Ascomycota: Hypocreales). Vet. Parasitol. 160(3-4):279-284. |
|
|
Niassy S, Maniania NK, Subramanian S, Gitonga LM, Ekesi S (2012a). Performance of a semiochemical-baited autoinoculation device treated with Metarhizium anisopliae for control of Frankliniella occidentalis on French bean in field cages. Entomol. Exp. Appl. 142(2):97-103. |
|
|
Niassy S, Maniania NK, Subramanian S, Gitonga ML, Maranga R, Obonyo AB, Ekesi S (2012b). Compatibility of Metarhizium anisopliae isolate ICIPE 69 with agrochemicals used in French bean production. J. Pest Manage. 58(2):131-137. |
|
|
Nyasani JO, Meyhöfer R, Subramanian S, Poehling H-M (2013). Seasonal abundance of western flower thrips and its natural enemies in different French bean agroecosystems in Kenya. J. Pest Sci. 86(3): 515-523. |
|
|
Ortiz-Ribbing L, Williams MM (2006). Conidial germination and germ tube elongation of Phomopsis amaranthicola and Microsphaeropsis amaranthi on leaf surfaces of seven Amaranthus species: Implications for biological control. Biol. Control 38(3):356-362. |
|
|
R Development Core Team (2014). A Language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.r-project.org/. |
|
|
Rieske LK, Raffa KF (2003). Evaluation of Visual and Olfactory Cues for Sampling Three Thrips Species (Thysanoptera: Thripidae) in Deciduous Forests of the Northern United States. J. Econ. Entomol. 96(3):777-782. |
|
|
Sampson C, Kirk WDJ (2013). Can mass trapping reduce thrips damage and is it economically viable? Management of the Western Flower Thrips in strawberry. PLoS ONE 8: e80787. |
|
|
Sidahmed OAA, Taha AK, Hassan GA, Abdalla IF (2014). Evaluation of pheromone dispenser units in methyl eugenol trap against Bactrocera invadens Drew, Tsuruta and White (Diptera: Tephritidae) in Sudan. J. Agric. Res. 3(8):148-151. |
|
|
Suckling DM, Walker JTS, Clare GK, Boyd Wilson KSH, Hall C, El-Sayed AM, Stevens PS (2012). Development and commercialisation of pheromone products in New Zealand. New Zealand Plant Prot. 65:267-273. |
|
|
Teulon DAJ, Casta-é C, Nielsen M-C, El-Sayed AM, Davidson MM, Gardner-Gee R, Poulton J, Kean AM, Hall C, Butler RC, Sansom CE, Suckling DM, Perry NB (2014). Evaluation of new volatile compounds as lures for western flower thrips and onion thrips in New Zealand and Spain. New Zealand Plant Prot. 67:175-183. |
|
|
Teulon DAJ, Davidson MM, Ducan IH, Dale EJ, Callum DF, Lesley L, Vanessa CG, Nigel BP (2007). 4-Pyridyl carbonyl and related compounds as thrips lures: effectiveness for onion thrips and New Zealand flower thrips in field experiments. J. Agric. Food Chem. 55(15):6198-6205. |
|
|
Teulon DAJ, Davidson MM, Nielsen M, Perry N, Van Tol R, de Kogel W (2010). The lure of scent: allelochemicals for thrips pest management. J. Insect Sci. 10(141):49-50. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


