Full Length Research Paper

ABSTRACT
INTRODUCTION
Maize is one of the most widely cultivated cereal crops in the world. In Sub-Saharan Africa, it is a staple food for both humans and animals. Unfortunately, this cereal belongs to the top two crops that are susceptible to fungal infection. During their growth, fungi may produce mycotoxins, not only in field conditions but also during storage. Mycotoxins are secondary metabolites produced by fungi, typically, species of Aspergillus, Fusarium or Penicillium. When inhaled, ingested, or absorbed through the skin, mycotoxins may cause adverse health effects for exposed humans and animals, such as lowered performance and sickness, and, at high concentrations, even death (Wagacha and Muthomi, 2008).
Thus, mould infection and mycotoxin production in grain are a health concern that negatively affect food and feed production, leading to environmental and socioeconomic consequences. A pre-requisite for the transition to more sustainable crop production in the future is the development of environmentally-friendly, efficient and cost-effective agricultural technology that minimises the growth of mould and spoilage of cereal crops during storage in developing countries. Innovative biocontrol methods may be part of this strategy, with the goal of protecting both animal and human health.
Yeasts, in general, are microorganisms which are highly resistant and adaptable to environmental change. Wickerhamomyces anomalus (formerly Hansenula anomala, Pichia anomala) J121 is able to grow within a wide range of temperatures (3-37ºC) and pH values (2-12), at low water activity (0.92 [NaCl] and 0.85 [glycerol]), as well as in anaerobic conditions. It can also assimilate a substantial number of different carbon and nitrogen sources (Fredlund et al., 2002). These characteristics bode well for the application of such a yeast in the context of maize storage on-farm.
Previous studies have shown that W. anomalus has beneficial protective effects on both food and feed, such as mould-inhibition. Production of ethyl acetate appears to be the mechanism of inhibition of mould growth, whereas the mechanism by which W. anomalus inhibits growth of Enterobacteriaceae is not fully understood (Olstorpe and Passoth, 2011). Yeasts can contribute to the value of the feed, because they contain substantial amounts of vitamins, minerals and proteins (Olstorpe et al., 2010). In addition, the degradation of inositol hexaphosphate (phytate) in cereal grain by yeast phosphatases releases phosphorus as well as chelated essential trace minerals (Sandberg and Andlid, 2002; Olstorpe et al., 2009).
The normal spectra of microbial populations present in traditionally cropped maize during air-dried storage in Cameroon have been investigated (Leong et al., 2012). Meyerozyma guilliermondii was the dominant yeast during extended storage (5 months), whilst W. anomalus was not found to be naturally present.
The aim of this study was to evaluate the efficacy of W. anomalus as a biocontrol yeast of moist maize grain post-harvest and during long-term airtight storage in Cameroon. A secondary aim was to monitor the feed hygiene of moist maize grain in such a storage system (yielding a type of fermentation), by enumeration and identification of different microbial groups.
MATERIALS AND METHODS
Yeast isolate
The yeast W. anomalus J121 (CBS 100487) used during the study was originally isolated from stored grain in Sweden. The yeast is stored in the fungal collection of the Department of Microbiology, Swedish University of Agricultural Sciences (SLU), Uppsala, Sweden, in glycerol stocks at -70°C.
Experimental design
The experiment was conducted at a farm in Nforya-Bamenda, Cameroon, at 1239 m above sea level, approximately latitude 6° north, and longitude 10° east. The white maize cultivar ‘Kasai’ was harvested at the beginning of August. Immediately after harvest, husks were removed and kernels shelled from the cobs by hand. Kernels were divided into two treatments: 39 kg of W. anomalus inoculated maize (‘inoculated’), and 39 kg of non-inoculated maize containing only the natural microbiota (‘control’). Both inoculated and control maize were further divided into triplicate portions, each comprising 13 kg of maize packed in black 20 L airtight plastic drums with removable lids. The control grain was harvested and packed first (that is, before the inoculated maize), to minimise the risk of cross-contamination between treatments. After packing, the drums were closed to generate airtight conditions. Samples were collected from each replicate at harvest (post-inoculation), and after 2, 5 and 8 months of storage.
Maize inoculation
To simulate commercial production of yeast inoculum, W. anomalus J121 was cultured in pilot-scale on molasses, ammonia, phosphorus, magnesium and vitamins by Jästbolaget (Rotebro, Sweden), dried with emulsifier on a fluidized bed, and vacuum-packed into 50 g portion sachets. Prior to inoculation, yeast was rehydrated with tap water. The yeast suspension was mixed by hand into the maize, calculated to inoculate 1×105 cells/g moist maize.
Analytical methods
The moisture content (MC) of the maize was determined by drying samples at 103°C for 16 h. Grain water activity (aw) was analysed using an AquaLab CX-2 (Decagon Devices Inc., Pullman, WA, USA) at 22°C.
Analysis of amino acids
The amino acid content of triplicate samples collected at harvest and after 8 months storage from both treatments was analysed by Eurofins AB, Lidköping, Sweden. Amino acid measurements were performed according to standard SS-EN ISO 13903:2005, except tryptophan that was analysed according to EU standard (EU Dir 2000/45/EG part C).
Quantification of microorganisms
Samples (20 g) from each triplicate were diluted with 180 ml sterile peptone water (Bacteriological peptone 2 g l−1; Merck, KGaA., Darmstadt, Germany), supplemented with 0.15 g l−1 Tween 80 (Merck, OHG., Schuchardt, Hohenbrunn, Germany), and homogenised for 120 s at normal speed in a Stomacher 400 Laboratory blender (Seward Medical, London, UK). Serial dilutions were performed in peptone water and spread onto various solid culture media. The LAB were quantified on de Man Rogosa Sharp (MRS) agar (Merck, KGaA., Darmstadt, Germany) supplemented with 100 µg/ml Delvocide (active compound, natamycin; Gist-Brocades B.V., Ma Delft , The Netherlands) to inhibit fungal growth. Plates were incubated anaerobically using a GasPack system (Becton Dickinson; Sparks, Md., USA) at 30°C for 48 h. Yeasts were enumerated on yeast peptone dextrose agar (YPD) at 25°C for 2-3 days, supplemented with 100 µg/ml chloramphenicol (Sigma-Aldrich Inc., St. Louis, MO) to inhibit bacterial growth. Moulds were quantified on YPD plates with a supplement of 100 µg/ml chloramphenicol and 10 µg/ml cycloheximide (Sigma-Aldrich Inc., St Louis, USA) to prevent growth of bacteria and yeasts, respectively, and the plates were incubated at 25°C for 3?4 days. Enterobacteriaceae were enumerated on Violet Red Bile Agar (Oxoid Ltd., Basingstoke, Hampshire, UK) by the pour plate method and incubated at 37°C for 24 h. Fungal and bacterial counts were expressed as Log10 mean (n=3) cfu/g feed.
DNA extraction, amplification and microbial identification
A total of 10 colonies of LAB and 10 colonies of yeast were randomly selected from each triplicate in both treatments at each sampling occasion. Colony PCR was performed for LAB isolates by selecting pure colonies as template. The 2 µl DNA template from yeast was extracted by boiling a single yeast colony in 50 µl nuclease-free water (Leong et al., 2012). PCR fingerprint patterns were generated by using the PuRe Taq ready-to-go PCR beads (GE Healthcare- Life Sciences, Uppsala Sweden) mixed with primer, DNA and water according to the instructions in the supplier’s manual. Genotypic differentiation was studied with repetitive-DNA-element PCR fingerprinting, using the microsatellite primer GTG (5’-GTGGTGGTGGTG GTG-3’). The rRNA gene amplification and PCR conditions for yeast and LAB fingerprints were performed as described by Olstorpe et al. (2008). The PCR fingerprints were visualised with electrophoresis on a 1% agarose gel in 0.5× Tris-borate-EDTA. The yeast and LAB fingerprints were analysed with GelCompar II version 4.5 software (Applied Maths, Kortrijk, Belgium). Strains representing each unique profile were identified by amplifying the 16S rRNA gene in LAB (primers 16Ss / 16Sr) or D1 / D2 region of the 26S rRNA in yeasts (primers NL1 / NL4). Amplicons were sequenced at Macrogen, Korea, and isolates were identified by sequence comparison against the Genbank database. Representative mould isolates were identified by a combination of morphological and molecular techniques. The morphological data and the DNA sequences were combined for polyphasic identification using Genbank and the CBS database as previously described (Leong et al., 2012).
Strain conservation of LAB and yeast
Yeast and LAB that were identified by sequence comparison were conserved and stored at the culture collection of the Department of Microbiology, Swedish University of Agricultural Sciences (SLU), Uppsala, Sweden. LAB were inoculated in 9 ml MRS-broth (Oxoid Ltd., Basingstoke, Hampshire, England) and incubated at 30°C overnight. Thereafter the tubes were centrifuged at 1000 g for 10 min and the supernatant discarded. The pellet was resuspended in 1 ml cryomedia (K2HPO4, 0,82 g/l; KH2PO4, 0.18 g/l; Na-citrate-dihydrate 0.67 g/l; MgSO4.7H2O, 0.25 g/l; 172 ml glycerol), and stored at –70°C. Yeast were inoculated in 2 ml yeast extract-peptone-D-glucose (YPD) broth (yeast extract, 10 g/l, bacteriological peptone, 20 g/l and D-glucose, 20 g/l) and incubated on a shaking board (130 rpm) at 30°C overnight. 1 ml of the suspension was mixed with an equal volume of glycerol and frozen at –70°C (Olstorpe et al., 2008).
Statistical analysis
The data generated from the nutritional analyses of the grain and the microbial counts were non-normally distributed; hence, a Wilcoxon ranking test suited for such non-parametric data was selected as the statistical analysis method. The effect of W. anomalus inoculation on nutritional values after storage was assessed using pairwise differences (harvest minus ‘after storage’) for each replicate as the input data. To estimate the effect of inoculation on microbial counts, the input data comprised counts obtained after 2, 5 and 8 months of storage. Mean values were considered significantly different at the 5% level.
RESULTS
Nutritional evaluation of the stored grain
After storage, the dry matter, crude protein, crude fibre, crude fats, and total amino acid contents did not differ significantly between the control maize and inoculated maize (P>0.05; Table 1). Of these parameters, dry matter, crude fibre and total amino acids had decreased from harvest levels to equivalent extents in both control and inoculated treatments. Inoculation with W. anomalus had significantly positive effects on the final contents of starch, and of three amino acids – alanine, aspartic acid and glycine compared with the control (P<0.05).
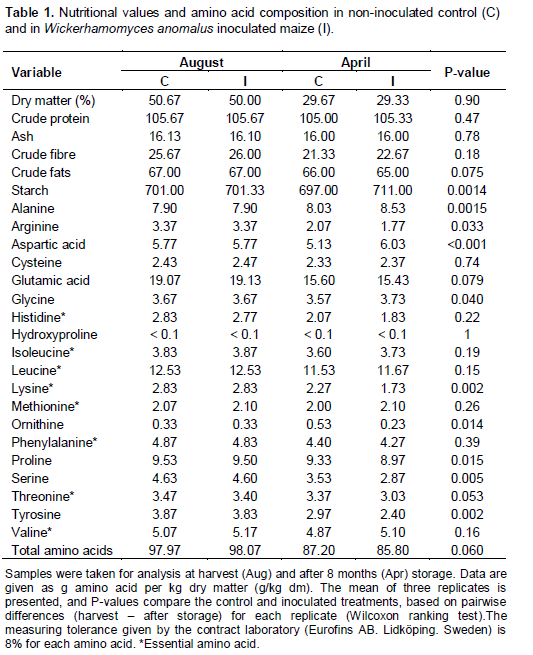
Starch and aspartic acid had decreased in the control samples during storage, whereas these were increased above harvest levels in the inoculated treatment. Alanine was increased after fermentation storage in both treatments, but to a greater extent in the inoculated samples. Glycine decreased in the control during storage, whereas it was maintained at harvest levels in the inoculated treatment. However, five amino acids decreased significantly more in inoculated than in control maize (P<0.05), namely, arginine, lysine, proline, serine and tyrosine; and ornithine was increased in the control maize but decreased in the inoculated maize after storage.
Microbial quantification
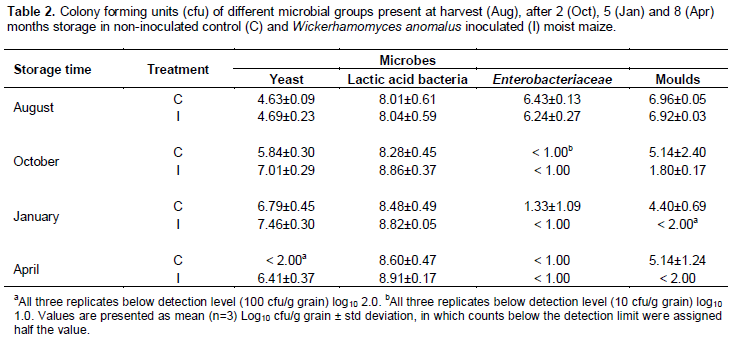
At harvest, microbial counts did not differ significantly between the control maize and the inoculated maize (Table 2). Yeast counts in both treatments increased up to 5 months, and then decreased markedly in the 8 month samples. Yeast counts increased more and maintained higher levels in the inoculated samples than in control maize, which by 8 months had reduced to levels below the detection limit. These overall elevated yeast counts in inoculated maize during storage were significantly higher than in control maize (P<0.001). Counts of LAB were fairly similar between treatments (P=0.02), maintaining levels similar to those present at harvest. Counts of Enterobacteriaceae decreased in both treatments after 2 months, and remained around the detection limit (log10 1 cfu/g) for the remainder of storage; thus, statistical comparison of inoculated and control samples during storage was not meaningful. Mould counts showed a similar pattern: decreasing within the first 2 months, and then remaining around those levels until end of storage. Counts in inoculated maize decreased to levels at or below the limit of detection after 2 months and remained at those levels, whereas the decrease in counts in control maize was less substantial; hence, overall counts during storage were significantly lower in inoculated maize than control maize (P<0.001).
Identified microbial species
Yeast
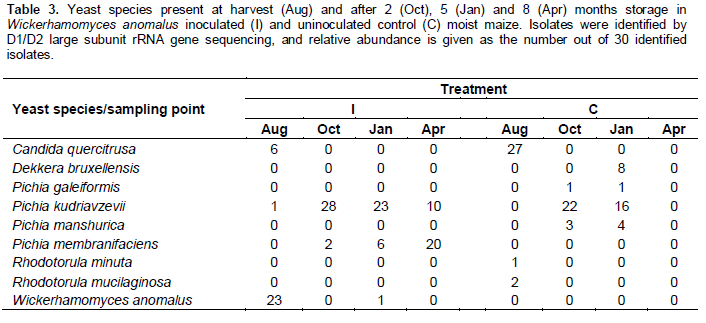
Candida quercitrusa was the dominant yeast isolated from the control maize at harvest, whereas, as expected, W. anomalus was dominant in the inoculated maize (Table 3). At 2 and 5 months, the dominant yeast species was Pichia kudriavzevii in both treatments. After 8 months, Pichia membranifaciens was the dominant yeast in the inoculated maize; yeasts were not isolated from the control maize.
Lactic acid bacteria
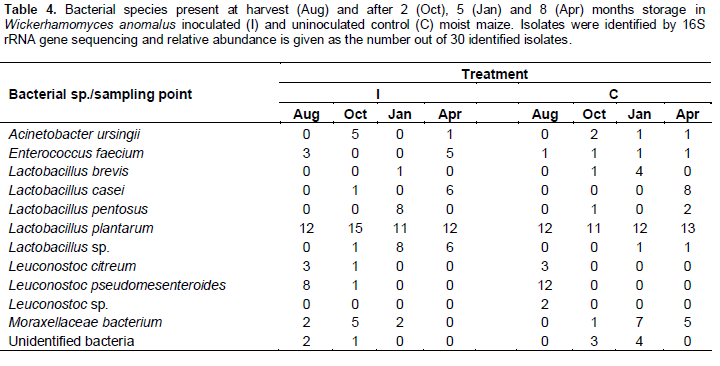
Changes in LAB populations during storage were fairly similar in control and inoculated maize (Table 4). Leuconostoc spp. and Lactobacillus plantarum were common at harvest; the latter, together with other species of Lactobacillus, continued to be frequently isolated throughout the storage period.
Moulds
Mould species isolated at harvest included Arthrinium arundinis, Aspergillus melleus, Bionectria ochroleuca, and Pleosporales sp. These species were not subsequently isolated during storage, in keeping with the overall decrease in mould counts (Table 2). Paecilomyces variotii was the dominant species isolated from maize during storage.
DISCUSSION
Previous studies have reported clear positive effects of W. anomalus inoculation, either as fresh yeast or freeze-dried preparations, on the amino acid and protein contents of moist stored grain, for example, in crimped barley stored in Swedish farm conditions for 5 months (Olstorpe, 2008; Olstorpe et al., 2010). Such effects were not observed in this first trial of moist (fermented) maize in Cameroon; rather, storage fermentation of maize with W. anomalus was not detrimental to the nutritional content, neither was it overtly positive. The total amino acids con-tent did not differ significantly between the control and inoculated maize after storage, in that both treatments showed a decrease compared with levels at harvest. In inoculated samples, the contents of three amino acids were improved or maintained at harvest levels, but contents of six other amino acids, including the essential amino acid, lysine, decreased more than in control maize. The absence of clear improvements during this maize storage trial, compared with previous reports from other grains, may be due to the loss of the biocontrol yeast W. anomalus after 2 months’ storage, with the population shifting towards P. kudriavzevii at 5 months, and P. membranifaciens at the end of storage. These latter species may not be associated with improved protein production for moist stored grain. Furthermore, yeast populations in inoculated maize were at their peak after 5 months of storage (log 7.46 cfu/g grain) and decreased to log 5.08 cfu/g at 8 months of storage. Protein evaluation was conducted at the end of the study when yeast numbers were lower, and no W. anomalus was present at that time.
In this study, the most notable finding in the use of W. anomalus for biopreservation of moist grain is the significant reduction in mould growth when the control and inoculated treatments were compared up to 8 months of storage. Note that the greatest reduction in mould counts was observed from harvest to 2 months’ storage, that is, after the initial inoculation with W. anomalus. Thus, the greater mould reduction in inoculated c.f. control maize could well be attributed to this biocontrol yeast, despite it apparently having died off in the 2 months and subsequent samples. The reduction in mould growth for inoculated maize has important implications for maize spoilage in the tropics, as the prevalence of moulds in stored grains is of public health significance due to risks for mycotoxin production (Wagacha and Muthomi, 2008). The main mould isolated during storage was the non-toxigenic species, Paecilomyces variotii, which is favoured by warm, dry conditions, with an optimal temperature of 35-40°C (Pitt and Hocking, 2009), and, thus, is a potential human pathogen in immunocompromised hosts. It is also thought to grow well under low oxygen tensions, which may be created during airtight storage of moist maize. This mould was eliminated in the inoculated maize after 8 months’ storage, which is a favourable outcome. Other studies have demonstrated the ability of W. anomalus to reduce mould growth through production of ethyl acetate during storage of crimped moist cereal grains and cereal by-products (Olstorpe and Passoth, 2011): for instance, in barley (Passoth et al., 2006; Olstorpe, 2008; Olstorpe et al., 2010), and wheat and oats (Petersson and Schnürer, 1998).
A further benefit of fermentation storage of moist maize grain demonstrated in this study was the reduction in Enterobacteriaceae to minimum detectable levels; this effect was observed in both control and inoculated grain, and is beneficial for grain hygiene and for reducing the risk of gastro-enteritis from pathogenic species that might be present. The constant levels of LAB observed in both treatments throughout storage and their production of organic acids and other anti-bacterial compounds may have contributed to the deleterious effect on Enterobacteriaceae (Lindgren and Dobrogosz, 1990). Similar reductions in Enterobacteriaceae during moist grain fermentations have been previously reported, particular with the addition of biocontrol yeast (Olstorpe et al., 2010; Olstorpe et al., 2012).
CONCLUSIONS
Moist storage (fermentation) of maize was sufficient and effective in reducing Enterobacteriaceae, a key parameter when monitoring grain hygiene; however, reducing the presence and growth of moulds during storage, thus minimising the risk for mycotoxin pro-duction, required inoculation with the biocontrol yeast, W. anomalus. The biocontrol yeast did not clearly improve the protein or amino acid contents of the stored maize in this trial, unlike previous trials with other grains. This could be attributed to its poor survival after 2 months; thus, formulation and preparation of the biocontrol ino-culum are avenues of further study, in order to optimise its survival and efficacy during moist maize storage in Cameroon.
CONFLICT OF INTERESTS
The author(s) have not declared any conflict of interests.
ACKNOWLEDGEMENTS
This project was supported by FORMAS (The Swedish Research Council for Environment, Agricultural Sciences and Spatial Planning) and the Swedish Research Council. We thank Mikael Andersson Franko, Department of Economics, SLU, for the statistical analyses.
REFERENCES
|
Fredlund E, Druvefors U, Boysen ME, Lingsten KJ, Schnürer J (2002). Physiological characteristics of the biocontrol yeast Pichia anomala J121. FEMS Yeast Res. 2:395-402. |
|
|
Leong SL, Niba AT, Ny S, Olstorpe M (2012). Microbial populations during maize storage in Cameroon. Afr. J. Biotechnol. 11: 8692-8697. |
|
|
Lindgren SE, Dobrogosz WJ (1990). Antagonistic activities of lactic acid bacteria in food and feed fermentations. FEMS Microbiol. Lett. 87:149-164. |
|
|
Olstorpe M (2008). Feed Grain Improvement through Biopreservation and Bioprocessing: Microbial Diversity, Energy Conservation and Animal Nutrition Aspects. PhD thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 77pp. |
|
|
Olstorpe M, Borling J, Schnürer J, Passoth V (2010). Pichia anomala yeast improves feed hygiene during storage of moist crimped barley grain under Swedish farm conditions. Anim. Feed Sci. Technol. 156:47-56. |
|
|
Olstorpe M, Lyberg K, Lindberg J-E, Schnürer J, Passoth V (2008). Population diversity of yeasts and lactic acid bacteria in pig feed fermented with whey, wet wheat distillers' grains or water at different temperatures. Appl. Environ. Microbiol. 74:1696-1703. |
|
|
Olstorpe M, Passoth V (2011). Pichia anomala in grain biopreservation. Antonie van Leeuwenhoek 99:57-62. |
|
|
Olstorpe M, Schnürer J, Passoth V (2009). Screening of yeast strains for phytase activity. Yeast Res. 9:478-488. |
|
|
Olstorpe M, Schnürer J, Passoth V (2012). Growth inhibition of various Enterobacteriaceae species by the yeast Hansenula anomala during storage of moist cereal grain. Appl. Environ. Microbiol. 78:29 |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


