Full Length Research Paper

ABSTRACT
INTRODUCTION
MATERIALS AND METHODS
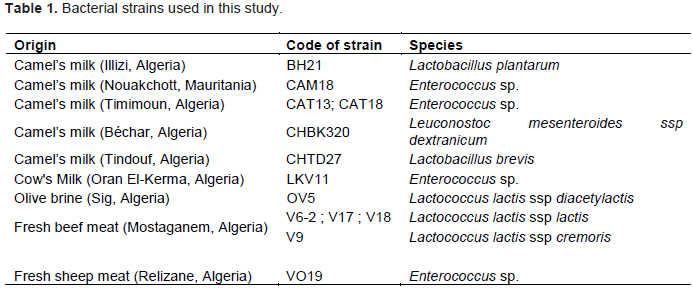
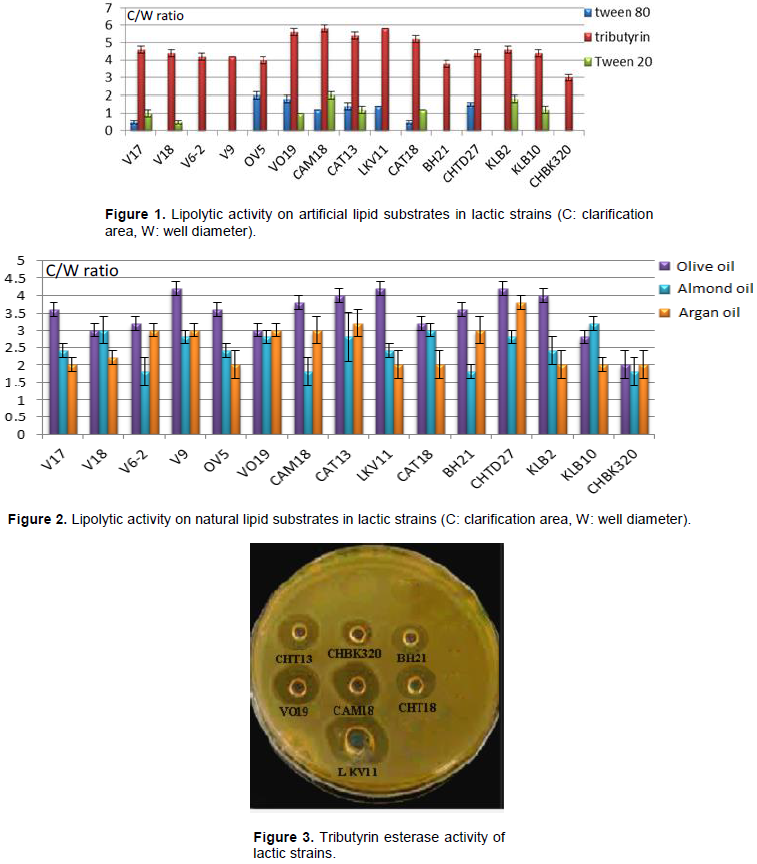
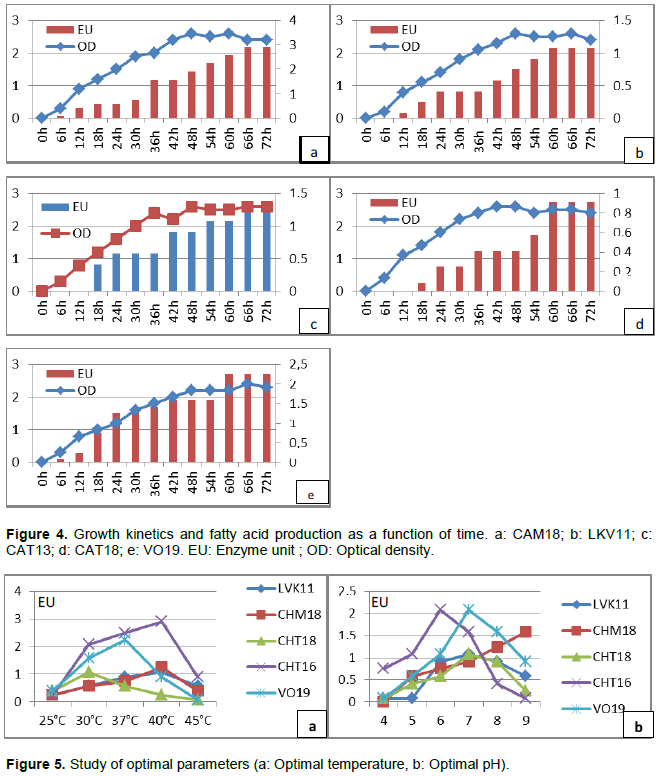
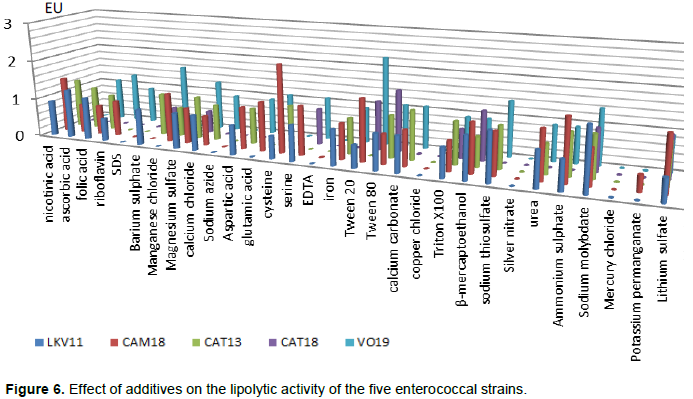
RESULTS AND DISCUSSION

CONCLUSION
CONFLICT OF INTERESTS
The authors declared no conflict of interests.
ACKNOWLEDGMENTS
REFERENCES
|
Andersen HJ, Ostdal H, Blom H (1995). Partial purification and characterisation of a lipase from Lactobacillus plantarum MF32. Food Chemistry 53:369-373. |
|
|
Bajaj A, Lohan P, Jha PN, Mehrotra R (2010). Biodiesel production through lipase catalyzed transesterification: an overview. Journal of Molecular Catalysis B Enzymatic 62:9-14. |
|
|
Borrelli GM, Trono D (2015). Recombinant lipases and phospholipases and their use as biocatalysts for industrial applications. International Journal of Molecular Sciences 16(9):20774-20840. |
|
|
Brennan NM, Ward AC, Beresford TP, Fox PF, Goodfellow M, Cogan TM (2002). Biodiversity of the bacterial flora on the surface of a smear cheese. Applied and Environmental Microbiology 68(2):820-830. |
|
|
Cardenas F, Alvarez E, Castro Alvarez MS, Sanchez- Montero JM, Valmaseda M, Elson SW, Sinisterra JV (2001). Screening and catalytic activity in organic synthesis of novel fungal and yeast lipases. Journal of Molecular Catalysis B Enzymatic 14:111-123. |
|
|
Castillo I, Requena T, Fernandez de Palencia P, Fontecha J, Gobbetti M (1999). Isolation and characterization of an intracellular esterase from Lactobacillus casei subsp.casei IFPL731. Journal of Applied Microbiology 86:653-659. |
|
|
Chich JF, Marchesseau K, Gripon JC (1997). Intracellular esterase from Lactococcus lactis subsp. Lactis NCDO 763: Purification and Characterization. International Dairy Journal 7:169- 174. |
|
|
Choi YJ, Miguez CB, Lee BH (2004). Characterization and Heterologous Gene Expression of a Novel Esterase from Lactobacillus casei CL96. Applied and Environmental Microbiology 70(6):3213-3221 |
|
|
Corrieu G, Luquet FM (2008). Bactéries lactiques - De la génétique aux ferments. Lavoisier, Paris, pp. 849. |
|
|
Dandavate V, Jinjala J, Keharia H, Madamwar D (2009). Production, partial purification and characterization of organic solvent tolerant lipase from Burkholderia multivorans V2 and its application for ester synthesis. Bioresources Technology 100(13):3374-3381. |
|
|
Das S, Holland R, Crow VL, Bennett RJ, Manderson GJ (2005). Effect of yeast and bacterial adjuncts on the CLA content and flavour of a washed-curd, dry-salted cheese. International Dairy Journal 15:807-815. |
|
|
De Man JC, Rogosa M, Sharpe ME (1960). A medium for the cultivation of lactobacilli. Journal of Applied Bacteriology 23(1):130-135. |
|
|
De Roissart H, Luquet FM (1994). Lactic acid bacteria, Vol. I et II, Edition Lorica, Uriage, France. |
|
|
Dharmsthiti S, Kuhasuntisuk B (1998). Lipase from Pseudomonas aeruginosa LP602: biochemical properties and application for wastewater treatment. Journal of Industrial Microbiology and Biotechnology 21:75-80. |
|
|
Dharmsthiti S, Luchai S (1999). Production, purification and characterization of thermophilic lipase from Bacillus sp. THL027. FEMS Microbiology Letters 179:241-246. |
|
|
Dharmsthiti S, Pratuangdejkul J, Theeragool GT, Luchai S (1998). Lipase activity and gene cloning of Acinetobacter calcoaceticus LP009. The Journal of General and Applied Microbiology 44:139-145. |
|
|
Dinçer E, Kıvanç M (2018). Lipolytic activity of lactic acid bacteria isolated from Turkish pastırma. Journal of Science and Technology C- Life Sciences and Biotechnology 7:1. |
|
|
El Soda M, Fathallah S, Ezzat N, Desmazeaud MJ, Abou Donia S (1986).The esterolytic and lipolytic activities of lactobacilli. Detection of the esterase systems of Lactobacillus casei, Lactobacillus plantarum, Lactobacillus brevis and Lactobacillus fermentum. Sciences des Aliments 6:545-547. |
|
|
Esakkiraj P, Rajikunar M, Palavesam A, Immanuel G (2010). Lipase production by Staphylococcus epidermidis CMSSTPI isolated from the gut of shrimp Penaeus indicus. Annals of Microbiology 60:37-42. |
|
|
Esteban-Torres M, Mancheño JM, De Las Rivas B, Muñoz R (2014b). Production and characterization of a tributyrin esterase from Lactobacillus plantarum suitable for cheese lipolysis. Journal of Dairy Science 97:6737-6744. |
|
|
Esteban-Torres M, Mancheño JM, De Las Rivas B, Muñoz R (2014a). Characterization of a halotolerant lipase from the lactic acid bacteria Lactobacillus plantarum useful in food fermentations. LWT - Food Science and Technology 60:246-252. |
|
|
Feitosa IC, Barbosa JMP, Orellana SC, Lima AS, Soares CMF (2010). Lipase production by microorganisms isolated from soils with a history of oil contact. Acta Scientiarum - Technology 32:27-31. |
|
|
Fenster KM, Parkin KL, Steele JL (2000). Characterization of an arylesterase from Lactobacillus helveticus CNRZ32. Journal of Applied Microbiology 88:572-583. |
|
|
Fernández L, Beerthuyzen MM, Brown J, Siizen RJ, Coolbear T, Holland R, Kuipers OP (2000). Cloning, characterization, controlled overexpression, and inactivation of the Major Tributyrin Esterase Gene of Lactococcus lactis. Applied and Environmental Microbiology 66(4):8-1360. |
|
|
Fickers P, Destain J, Thonart P (2008). Lipases are atypical hydrolases: main characteristics and applications. Biotechnologie, Agronomie, Société et Environnement 12:119-130. |
|
|
García-Cano I, Rocha-Mendoza D, Ortega-Anaya J, Wang K, Kosmerl E, Jiménez-Flores R (2019). Lactic acid bacteria isolated from dairy products as potential producers of lipolytic, proteolytic and antibacterial proteins. Applied Microbiology and Biotechnology 103:5243-5257. |
|
|
Ginalska G, Bancerz R, Korniffowicz-Kowalska T (2004). A thermostable lipase produced by a newly isolated Geotrichum-like strain, R59. Journal of Industrial Microbiology and Biotechnology 31:177-182. |
|
|
Gobbetti M, Fox PF, Smacchi E, Stepaniak L, Damiani P (1996). Purification and characterisation of a lipase from Lactobacillus plantarum 2739. Journal of Food Biochemistry 20:227-246. |
|
|
Gobbetti M, Fox PF, Stepaniak L (1997b). Isolation and characterisation of a tributyrin esterase from Lactobacillus plantarum 2739. Journal of Dairy Science 80:3099-3106. |
|
|
Gobbetti M, Smacchi E, Corsetti A (1997a). Purification and characterisation of a cell surface-associated esterase from Lactobacillus fermentum DT41. International Dairy Journal 7:13-21. |
|
|
Guiraud JY, Galzy P (1980). Microbiological analysis in the food industries. Edition de l'usine, p 39. |
|
|
Gupta R, Gupta N, Rathi P (2004). Bacterial lipases: an overview of production, purification and biochemical properties. Applied Microbiology and Biotechnology 64 (6):763-781. |
|
|
Holland R, Coolbear T (1996). Purification of Tributyrin Esterase from Lactococcus lactis subsp cremoris E8. Journal of Dairy Research 63:131-140. |
|
|
Hong Q, Liu XM, Hang F, Zhao JX, Zhang H, Chen W (2018). Screening of adjunct cultures and their application in ester formation in Camembert-type cheese. Food Microbiology 70:33-41. |
|
|
Iqbal AS, Rehman A (2015). Characterization of lipase from Bacillus subtilis I-4 and its potential use in oil contaminated wastewater. Brazilian Archives of Biology and Technology 58(5):789-797. |
|
|
Joseph B, Ramteke PW, Thomas G (2008). Cold active microbial lipases: some hot issues and recent developments. Biotechnology Advances 26:457-470. |
|
|
Kanwar L, Goswami P (2002). Isolation of a Pseudomonas lipase produced in pure hydrocarbon substrate and its applications in the synthesis of isoamyl acetate using membrane-immobilized lipase. Enzyme and Microbial Technology 31:727-735. |
|
|
Katz M, Medina R, Gonzalez S, Oliver G (1997). Esterolytic and Lipolytic Activities of Lactic Acid Bacteria Isolated from Ewes Milk and Cheese. Journal of Food Protection 65:2002-2001. |
|
|
Khalid NM, El Soda M, Marth EH (1990). Esterases of Lactobacillus helveticus and Lactobacillus delbrueckii spp.bulgaricus. Journal of Dairy Science 73:2711-2719. |
|
|
Kilcawley KN, Wilkinson M, Fox PF (1998). Enzyme modified cheese. International Dairy Journal 8:1-10. |
|
|
Kuhl GC, De J, Lindner D, Barron F (2016). Biohydrogenation of linoleic acid by lactic acid bacteria for the production of functional cultured dairy products: a review. Foods 5(1):13. |
|
|
Kumar D, Kumar L, Nagar S, Raina C, Parshad R, Gupta VK (2012). Screening, isolation and production of lipase/esterase producing Bacillus sp. strain DVL2 and its potential evaluation in esterification and resolution reactions. Archives of Applied Science Research 4:1763-1770. |
|
|
Lee OW, Koh YS, Kim KJ, Kim BC, Choi HJ, Kim DS, Suhartono MT, Pyun YR (1999). Isolation and characterization of a thermophilic lipase from Bacillus thermoleovorans ID-1. FEMS Microbiology Letters 179:393-400. |
|
|
Lesuisse E, Schanck K, Colson C (1993). Purification and preliminary characterization of the extracellular lipase of Bacillus subtilis 168, an extremely basic pH-tolerant enzyme. European Journal of Biochemistry 216:155-160. |
|
|
Litthauer D, Ginster A, Skein E (2002). Pseudomonas luteola lipase: a new member of the 320-residue Pseudomonas lipase family. Enzyme and Microbial Technology 30:209-215 |
|
|
Liu SQ, Holland R, Crow VL (2001). Purification and properties of intracellular esterases from Streptococcus thermophilus. International Dairy Journal 11:27-35. |
|
|
Lopes MF, Leitao AL, Regalla M, Figueiredo Marques JJ, Teixeira Carrondo MJ, Barreto Crespo MT (2002). Characterization of a highly thermostable extracellular lipase from Lactobacillus plantarum. International Journal of Food Microbiology 76:107-115 |
|
|
Makhzoum A, Knapp JS, Owusu RK (1995). Factor affecting growth and lipase production by Pseudomonas fluorescens 2D. Food Microbiology 12:277-290 |
|
|
Martinelle M, Holmquist M, Hult K (1995). On the interfacial activation of Candida antarctica lipase A and B as compared with Humicola lanuginosa lipase. Biochimica et Biophysica Acta (BBA) - Lipids and Lipid Metabolism 1258:272-6. |
|
|
Medina RB, Katz MB, González S, Oliver G (2004). Methods to Determination of Esterolytic and Lipolytic Activities of Lactic Acid Bacteria. Methods in molecular biology, public Health microbiology. Methods and protocols. John F. T. Spencer (Editor), Alicia L. Ragout de Spencer (Series Editor) pp:437-441. |
|
|
Meyers SA, Cuppett SL, Hutkins RW (1996). Lipase Production by |
|
|
Lactic Acid Bacteria and Activity on Butter Oil. Food Microbiology 13(5):383-389. |
|
|
Mobarak-Qamsari E, Kasra-Kermanshahi R, Moosavi-nejad Z (2011). Isolation and identification of a novel, lipase-producing bacterium, Pseudomnas aeruginosa KM110. Iranian Journal of Microbiology 3(2):92-98. |
|
|
Nardi M, Fiez-Vandal C, Tailliez P, Monnet V (2002). The EstA esterase is responsible for the main capacity of Lactococcus lactis to synthesize short chain fatty acid esters in vitro. Journal of Applied Microbiology 93:994-1002. |
|
|
Nwachukwu E, Ejike EN, Ejike BU, Onyeanula EO, Chikezie-abba RO, Okorocha NA, Onukaogu UE (2017). Characterization and optimization of lipase production from soil microorganism (Serratia marcescens). International Journal of Current Microbiology and Applied Sciences 6(12):1215-1231. |
|
|
Palanichamy E, Rajamony U, Arunachalam P, Grasian I (2012). Solid-state production of esterase using fish processing wastes by Bacillus altitudinis AP-MSU. Food and Bioproducts Processing 90:370-376. |
|
|
Papon M, Talon R (1988). Factors affecting growth and lipase production by meat lactobacilli strains and Brochothrix thermosphacta. Journal of Applied Bacteriology 64:107-115 |
|
|
Ramakrishnan V, Goveas LC, Suralikerimath N, Jampani C, Halami PM, Narayan B (2016). Extraction and purification of lipase from Enterococcus faecium MTCC5695 by PEG/phosphate aqueous-two phase system (ATPS) and its biochemical characterization. Biocatalysis and Agricultural Biotechnology 6:19-27. |
|
|
Salwoom L, Raja Abd Rahman RNZ, Salleh AB, Mohd Shariff F, Convey P, Pearce D, Mohamad Ali MS (2019). Isolation, Characterisation, and Lipase Production of a Cold-Adapted Bacterial Strain Pseudomonas sp. LSK25 Isolated from Signy Island, Antarctica. Molecules 24(4):715. |
|
|
Saxena P, Whang I, Lee J, Voziyanov Y, Mendoza V, Jayaram M (1994). Role of tyrosine phosphorylation-dephosphorylation in copy number control of the yeast plasmid 2 micron circle. Cellular and Molecular Biology Research 40(3):215-22. |
|
|
Schmidt-Dannert C, Sztajer H, Stocklein W, Menge U, Schmid RD (1994). Screening purification and properties of a thermophiliclipase from Bacillus thermocatenulatus. Biochimica et Biophysica Acta (BBA) - Lipids and Lipid Metabolism 1214:43-53 |
|
|
Serio A, Chaves-López C, Paparella A, Suzzi G (2010). Evaluation of metabolic activities of enterococci isolated from Pecorino Abruzzese cheese. International Dairy Journal 20:459-464. |
|
|
Sharma S, Kanwar K, Kanwar SS (2016). Ascorbyl palmitate synthesis in an organic solvent system using a Celite-immobilized commercial lipase (Lipolase 100L). 3 Biotech 6:183. |
|
|
Sidhu P, Sharma R, Soni SK, Gupta JK (1998a). Effect of cultural conditions on extracellular alkaline lipase production from Bacillus sp. RS-12 and its characterization. Indian Journal of Microbiology 38:9-14. |
|
|
Sidhu P, Sharma R, Soni SK, Gupta JK (1998a). Production of extracellular alkaline lipase by a new thermophillic Bacillus sp. Folia Microbiologica 43:51-54. |
|
|
Sokolovska I, Albasi C, Riba JP, Bales V (1998). Production of extracellular lipase by Candida cylindracea CBS 6330. Bioprocess Engineering 19:179-186. |
|
|
Soleymani S, Alizadeh H, Mohammadian H, Rabbani E, Moazen F, Sadeghi HM, Shariat ZS, Etemadifar Z, Rabbani M (2017). Efficient Media for High Lipase Production: One Variable at a Time Approach. Avicenna Journal of Medical Biotechnology 9(2):82-86. |
|
|
Sooch BS, Kauldhar BS (2013). Influence of multiple bioprocess parameters on production of lipase from Pseudomonas sp. BWS-5. Brazilian Archives of Biology and Technology 56(5):711-721. |
|
|
Sunna A, Hunter L, Hutton CA, Bergquist PL (2002). Biochemical characterization of a recombinant thermoalkalophilic lipase and assessment of its substrate enantioselectivity. Enzyme and Microbial Technology 31:472-476. |
|
|
Sztajer H, Lunsdorf H, Erdmann H, Menge U, Schmid R (1992). Purification and properties of lipase from Penicillium simplicissimum. Biochimica et Biophysica Acta (BBA) - Lipids and Lipid Metabolism 1124(3):253- 261 |
|
|
Talon R, Montel MC (1994). Activités estérasiques et lipolytiques des bactéries lactiques. in: Luquet FM, De Roissart H, Bacteries lactiques, Vol. 1, editions Coquand, Grenoble, France, pp. 349-352. |
|
|
Thirunavukarasu K, Edwinoliver NG, Anbarasan S, Gowthaman MK, Iefuji H, Kamini NR (2008). Removal of triglyceride soil from fabrics by a novel lipase from Cryptococcus sp.S-2. Process Biochemistry 43:701-6. |
|
|
Tsakalidou E, Dalezios I, Kalantzopoulos G (1994). Isolation and partial characterization of an intracellular esterase from Enterococcus faecium ACA-DC 237. Journal of Biotechnology 37:201-208. |
|
|
Vishnupriya B, Sundaramoorthi C, Kalaivani M, Selvam K (2010). Production of lipase from Streptomyces griseus and evaluation of bioparameters. International Journal of Chemtech Research 2(3):1380-1383. |
|
|
Wang Y, Srivastava KC, Shen GJ, Wang HY (1995). Thermostable alkaline lipase from a newly isolated thermophilic Bacillus, strain A30-1 (ATCC 53841). Journal of Fermentation and Bioengineering 79:433-438. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


