Review

ABSTRACT
Improved agricultural production of essential crops through advanced breeding is important for increasing access to nutritious food for the world's rapidly growing population, which is expected to reach 9.8 billion by 2050. Recent advancements in the clustered regularly interspaced short palindromic repeats/CRISPR-associated protein9 (CRISPR/Cas9) genome editing process, which uses single-guide RNA for genome editing, have made it easy, stable and efficient tool for targeted gene mutations, knockout and knock-in/replacement to boost crop yield. The CRISPR/Cas method is constantly being improved, and its applications have greatly expanded. It can be used to modify the genome sequence of any organism, including plants like cassava, to achieve the desired trait. As a result, CRISPR/Cas is regarded as a game-changing technology in plant biology. Here, we discuss the principles of operation, implementations and future prospects of CRISPR/Cas9 for efficient processing of individual genes in cassava cultures. Recent work on cassava crop with regards to the use of CRISPR/Cas9 for the plant improvement was also addressed.
Key words: Manihot esculenta, CRISPR/Cas9, genome editing, gRNA, protospacer adjacent motif (PAM).
INTRODUCTION
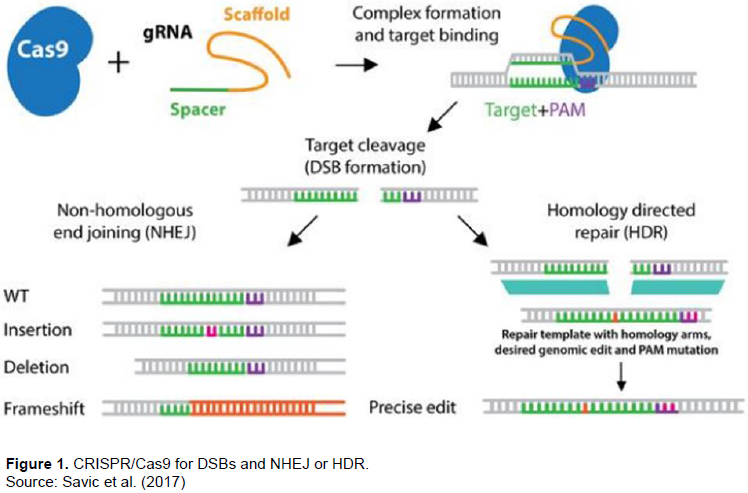
UNDERSTANDING CRISPR/CAS GENOME EDITING
MECHANISM OF CRISPR/CAS GENOME EDITING
DELIVERY OF CRISPR/CAS REAGENTS TO CASSAVA
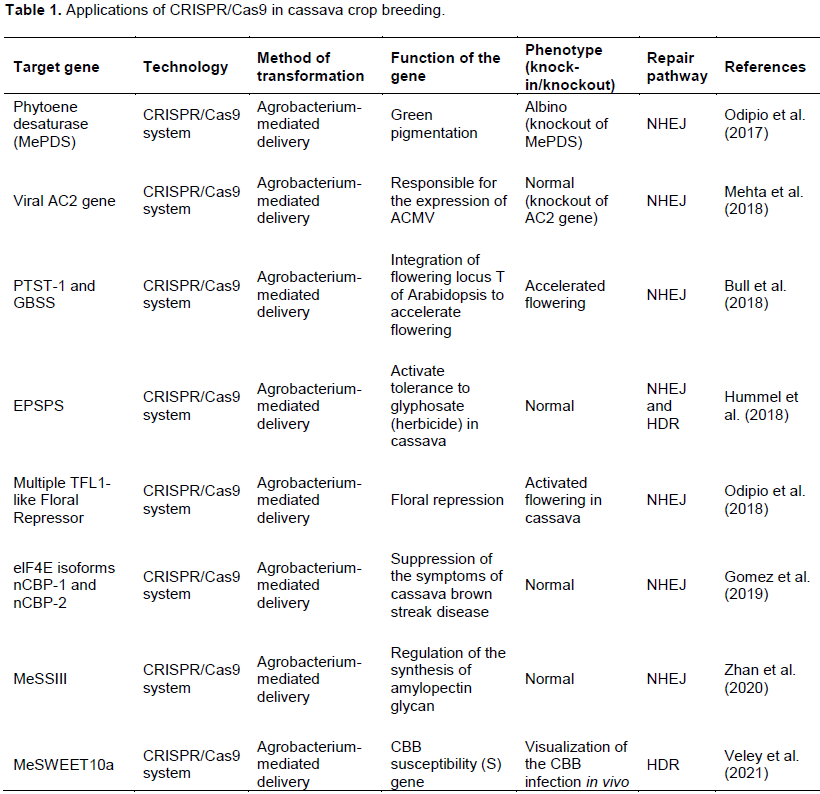
FUTURE PERSPECTIVES FOR CRISPR/CAS9 IN CASSAVA BREEDING AND RESEARCH
CONCLUSION AND RECOMMENDATIONS
CONFLICT OF INTERESTS
REFERENCES
|
Abdelrahman M, Al-Sadi AM, Pour-Aboughadareh A, Burritt DJ, Tran LSP (2018). Genome editing using CRISPR/Cas9-targeted mutagenesis: An opportunity for yield improvements of crop plants grown under environmental stresses. Plant Physiology and Biochemistry 131:31-36. |
|
|
Adhikari P, Poudel M (2020). CRISPR-Cas9 in agriculture: Approaches, applications, future perspectives, and associated challenges. Malaysian Journal of Halal Research 3(1):6-16. |
|
|
Al-Khayri JM, Jain SM, Johnson DV (2015). Advances in plant breeding strategies: Breeding, biotechnology and molecular tools. Advances in Plant Breeding Strategies: Breeding, Biotechnology and Molecular Tools (Vol. 1). |
|
|
Andersson M, Turesson H, Olsson N, Fält AS, Ohlsson P, Gonzalez MN, Hofvander P (2018). Genome editing in potato via CRISPR-Cas9 ribonucleoprotein delivery. Physiologia Plantarum 164(4):378-384. |
|
|
Ansari A, Wang C, Wang J, Wang F, Liu P, Gao Y, Zhao K (2017). Engineered dwarf male-sterile rice: A promising genetic tool for facilitating recurrent selection in rice. Frontiers in Plant Science 8:1-11. |
|
|
Arora L, Narula A (2017). Gene editing and crop improvement using CRISPR-cas9 system. Frontiers in Plant Science 8:1932. |
|
|
Barrangou R, Horvath P (2017). A decade of discovery: CRISPR functions and applications. Nature Microbiology 2(7):1-9. |
|
|
Bernheim A, Calvo-Villamañán A, Basier C, Cui L, Rocha EPC, Touchon M, Bikard D (2017). Inhibition of NHEJ repair by type II-A CRISPR-Cas systems in bacteria. Nature Communications 8(1):25-28. |
|
|
Bhatta BP, Malla S (2020). Improving horticultural crops via crispr/cas9: Current successes and prospects. Plants 9(10):1-19. |
|
|
Bhowmik P, Ellison E, Polley B, Bollina V, Kulkarni M, Ghanbarnia K, Kagale S (2018). Targeted mutagenesis in wheat microspores using CRISPR/Cas9. Scientific Reports 8(1):1-10. |
|
|
Braatz J, Harloff HJ, Mascher M, Stein N, Himmelbach A, Jung C (2017). CRISPR-Cas9 targeted mutagenesis leads to simultaneous modification of different homoeologous gene copies in polyploid oilseed rape (Brassica napus). Plant Physiology 174(2):935-942. |
|
|
Bredeson JV, Lyons JB, Prochnik SE, Wu GA, Ha CM, Edsinger-Gonzales E, Rokhsar DS (2016). Sequencing wild and cultivated cassava and related species reveals extensive interspecific hybridization and genetic diversity. Nature Biotechnology 34(5):562-570. |
|
|
Bull SE, Seung D, Chanez C, Mehta D, Kuon JE, Truernit E, Vanderschuren H (2018). Accelerated ex situ breeding of GBSS- and PTST1-edited cassava for modified starch. Science Advances 4(9): eaat6086. |
|
|
Carter J, Wiedenheft B (2015). Snapshot: CRISPR-RNA-guided adaptive immune systems. Cell 163(1):260-260.e1. |
|
|
Ceballos H, Jaramillo J, Salazar S, Pineda L, Calle F, Setter T (2017). Induction of flowering in cassava through grafting. Journal of Plant Breeding and Crop Science 9(2)19-29. |
|
|
Char SN, Wei J, Mu Q, Li X, Zhang ZJ, Yu J, Yang B (2020). An Agrobacterium-delivered CRISPR/Cas9 system for targeted mutagenesis in sorghum. Plant Biotechnology Journal 18(2):319-321. |
|
|
Che P, Anand A, Wu E, Sander JD, Simon MK, Zhu W, Jones TJ (2018). Developing a flexible, high-efficiency Agrobacterium-mediated sorghum transformation system with broad application. Plant Biotechnology Journal 16(7):1388-1395. |
|
|
Chiurugwi T, Kemp S, Powell W, Hickey LT (2019). Speed breeding orphan crops. Theoretical and Applied Genetics 132(3):607-616. |
|
|
Christian M, Cermak T, Doyle EL, Schmidt C, Zhang F, Hummel A, Voytas DF (2010). Targeting DNA double-strand breaks with TAL effector nucleases. Genetics 186(2):756-761. |
|
|
Clarke JL, Zhang P (2013). Plant biotechnology for food security and bioeconomy. Plant Molecular Biology 83(1-2):1-3. |
|
|
Concordet JP, Haeussler M (2018). CRISPOR: Intuitive guide selection for CRISPR/Cas9 genome editing experiments and screens. Nucleic Acids Research 46(W1):W242-W245. |
|
|
Devkota S (2018). The road less traveled: Strategies to enhance the frequency of homology-directed repair (HDR) for increased efficiency of CRISPR/Cas-mediated transgenesis. BMB Reports 51(9):437-443. |
|
|
Doll NM, Gilles LM, Gérentes MF, Richard C, Just J, Fierlej Y, Widiez T (2019). Single and multiple gene knockouts by CRISPR-Cas9 in maize. Plant Cell Reports 38(4):487-501. |
|
|
Doudna JA, Charpentier E (2014). The new frontier of genome engineering with CRISPR-Cas9. Science 346(6213). |
|
|
Fauser F, Schiml S, Puchta H (2014). Both CRISPR/Cas-based nucleases and nickases can be used efficiently for genome engineering in Arabidopsis thaliana. Plant Journal 79(2):348-359. |
|
|
Gaj T, Gersbach CA, Barbas CF (2013). ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends in Biotechnology 31(7)397-405 |
|
|
Gao J, Wang G, Ma S, Xie X, Wu X, Zhang X, Xia Q (2015). CRISPR/Cas9-mediated targeted mutagenesis in Nicotiana tabacum. Plant Molecular Biology 87(1-2):99-110. |
|
|
Garrett RA, Vestergaard G, Shah SA (2011). Archaeal CRISPR-based immune systems: Exchangeable functional modules. Trends in Microbiology 19(11):549-556. |
|
|
Georges F, Ray H (2017). Genome editing of crops: A renewed opportunity for food security. GM Crops and Food 8(1):1-12. |
|
|
Ghogare R, Williamson-Benavides B, Ramírez-Torres F, Dhingra A (2020). CRISPR-associated nucleases: the Dawn of a new age of efficient crop improvement. Transgenic Research 29:1-35. |
|
|
Ghosh S, Watson A, Gonzalez-Navarro OE, Ramirez-Gonzalez RH, Yanes L, Mendoza-Suárez, M, Hickey LT (2018a). Speed breeding in growth chambers and glasshouses for crop breeding and model plant research. Nature Protocols 13(12):2944-2963. |
|
|
Ghosh S, Watson A, Gonzalez-Navarro OE, Ramirez-Gonzalez RH, Yanes L, Mendoza-Suárez, M, Hickey LT (2018b). Speed breeding in growth chambers and glasshouses for crop breeding and model plant research. Nature Protocols 13(12):2944-2963 BioRxiv. |
|
|
Godfray HCJ, Beddington JR, Crute IR, Haddad L, Lawrence D, Muir JF, Toulmin C (2010). Food security: The challenge of feeding 9 billion people. Science 327(5967):812-818. |
|
|
Gomez MA, Lin ZD, Moll T, Chauhan RD, Hayden L, Renninger K, Bart RS (2019). Simultaneous CRISPR/Cas9-mediated editing of cassava eIF4E isoforms nCBP-1 and nCBP-2 reduces cassava brown streak disease symptom severity and incidence. Plant Biotechnology Journal 17(2):421-434. |
|
|
Gupta RM, Musunuru K (2014). The emergence of genome-editing technology. The Journal of Clinical Investigation 124(10):4154-4161. |
|
|
Haque E, Taniguchi H, Hassan MM, Bhowmik P, Karim M R, ?miech M, Islam T (2018). Application of CRISPR/Cas9 genome editing technology for the improvement of crops cultivated in tropical climates: Recent progress, prospects, and challenges. Frontiers in Plant Science 9:617. |
|
|
Hatem A, Bozda? D, Çatalyürek ÜV (2011). Benchmarking short sequence mapping tools. Proceedings - 2011 IEEE International Conference on Bioinformatics and Biomedicine, BIBM 2011, 109-113. |
|
|
He Y, Zhu M, Wang L, Wu J, Wang Q, Wang R, Zhao Y (2018). Programmed Self-Elimination of the CRISPR/Cas9 Construct Greatly Accelerates the Isolation of Edited and Transgene-Free Rice Plants. Molecular Plant 11(9):1210-1213. |
|
|
Hoffmann MD, Aschenbrenner S, Grosse S, Rapti K, Domenger C, Fakhiri J, Niopek D (2019). Cell-specific CRISPR-Cas9 activation by microRNA-dependent expression of anti-CRISPR proteins. Nucleic Acids Research 47(13):e75. |
|
|
Hui-Li X, D L, Wang Z, Zhang H, Han C, Liu B, Chen Q (2014). A CRISPR/Cas9 toolkit for multiplex genome editing in plants. Plant Biology 1(1):3-13. |
|
|
Hummel AW, Chauhan RD, Cermak T, Mutka AM, Vijayaraghavan A, Boyher A, Taylor NJ (2018). Allele exchange at the EPSPS locus confers glyphosate tolerance in cassava. Plant Biotechnology Journal 16(7):1275-1282. |
|
|
Hussain B, Lucas SJ, Budak H (2018). CRISPR/Cas9 in plants: at play in the genome and at work for crop improvement. Briefings in Functional Genomics 17(5):319-328. |
|
|
Ishino Y, Shinagawa H, Makino K, Amemura M, Nakatura A (1987). Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isoenzyme conversion in Escherichia coli, and identification of the gene product. Journal of Bacteriology 169(12):5429-5433. |
|
|
Ishino, Yoshizumi, Krupovic, M, Forterre P (2018). History of CRISPR-Cas from Encounter with a Mysterious. Journal of Bacteriology 200(7):e00580-17. |
|
|
Jansen R, Van Embden JDA, Gaastra W, Schouls LM (2002). Identification of genes that are associated with DNA repeats in prokaryotes. Molecular Microbiology 43(6):1565-1575. |
|
|
Jasin M, Rothstein R (2013). Repair of strand breaks by homologous recombination. Cold Spring Harbor Perspectives in Biology 5(11):1-18. |
|
|
Jayavaradhan R, Pillis DM, Goodman M, Zhang F, Zhang Y, Andreassen PR, Malik P (2019). CRISPR-Cas9 fusion to dominant-negative 53BP1 enhances HDR and inhibits NHEJ specifically at Cas9 target sites. Nature Communications 10(1):1-13. |
|
|
Jinek M, Chylinski K, Fonfara I, Hauer M, Doudna JA, Charpentier E (2012). A Programmable Dual-RNA - Guided 337(6096):816-821. |
|
|
Jinek M, Jiang F, Taylor DW, Sternberg SH, Kaya E, Ma E, Doudna JA (2014). Structures of Cas9 endonucleases reveal RNA-mediated conformational activation. Science 343(6176):2-18. |
|
|
Khoury LYE, Campbell JM, Clark KJ (2018). The transition of zebrafish functional genetics from random mutagenesis to targeted integration. Molecular-Genetic and Statistical Techniques for Behavioral and Neural Research. Elsevier Inc. |
|
|
Kivrak E, Pauzaite T, Copeland NA, Hardy JG, Kara P, Firlak M, Ozsoz M (2021). Detection of CRISPR-Cas9-Mediated Mutations Using a Carbon Nanotube-Modified Electrochemical Genosensor. Biosensors 11(1):17. |
|
|
Labun K, Montague TG, Krause M, Torres Cleuren YN, Tjeldnes H, Valen E (2019). CHOPCHOP v3: Expanding the CRISPR web toolbox beyond genome editing. Nucleic Acids Research 47(W1):W171-W174. |
|
|
Lawrenson T, Shorinola O, Stacey N, Li C, Østergaard L, Patron N, Harwood W (2015). Induction of targeted, heritable mutations in barley and Brassica oleracea using RNA-guided Cas9 nuclease. Genome Biology 16(1):1-13. |
|
|
Lee K, Zhang Y, Kleinstiver BP, Guo JA, Aryee MJ, Miller J, Wang K (2019). Activities and specificities of CRISPR/Cas9 and Cas12a nucleases for targeted mutagenesis in maize. Plant Biotechnology Journal 17(2):362-372. |
|
|
Li C, Chen C, Chen H, Wang S, Chen X, Cui Y (2018). Verification of DNA motifs in Arabidopsis using CRISPR/Cas9-mediated mutagenesis. Plant Biotechnology Journal 16(8):1446-1451. |
|
|
Li G, Zhang X, Zhong C, Mo J, Quan R, Yang J, Wu Z (2017). Small molecules enhance CRISPR/Cas9-mediated homology-directed genome editing in primary cells. Scientific Reports 7(1):1-11. |
|
|
Li J, Manghwar H, Sun L, Wang P, Wang G, Sheng H, Zhang X (2019). Whole genome sequencing reveals rare off-target mutations and considerable inherent genetic or/and somaclonal variations in CRISPR/Cas9-edited cotton plants. Plant Biotechnology Journal, 17(5):858-868. |
|
|
Li Q, Sapkota M, van der Knaap E (2020). Perspectives of CRISPR/Cas-mediated cis-engineering in horticulture: unlocking the neglected potential for crop improvement. Horticulture Research 7(1). |
|
|
Li S, Xia L (2020). Precise gene replacement in plants through CRISPR/Cas genome editing technology: current status and future perspectives. ABIOTECH 1(1):58-73. |
|
|
Li T, Liu B, Spalding MH, Weeks DP, Yang B (2012). High-efficiency TALEN-based gene editing produces disease-resistant rice. Nature Biotechnology 30(5):390-392. |
|
|
Li, Zhan, Wang, Y, Lu X, Li, R, Liu J, Fu S, Yao Y (2020). Construction and Verification of CRISPR/Cas9 Gene Editing Vector for Cassava MeSSIII Gene. Molecular Plant Breeding 11(17):1-8. |
|
|
Li, Zhongsen, Liu, Z. Bin, Xing, A., Moon, B. P., Koellhoffer, J. P., Huang, L, Cigan, A. M. (2015). Cas9-guide RNA directed genome editing in soybean. Plant Physiology 169(2):960-970. |
|
|
Liu H, Ding Y, Zhou Y, Jin W, Xie K, Chen LL (2017). CRISPR-P 2.0: An Improved CRISPR-Cas9 Tool for Genome Editing in Plants. Molecular Plant 10(3):530-532. |
|
|
Liu Q, Chen B, Wang Q, Shi X, Xiao Z, Lin J, Fang X (2009). Carbon nanotubes as molecular transporters for walled plant cells. Nano Letters 9(3):1007-1010. |
|
|
Liu, Xuan, Wu, S, Xu J, Sui C, Wei J (2017). Application of CRISPR/Cas9 in plant biology. Acta Pharmaceutica Sinica B 7(3):292-302. |
|
|
Liu, Xuejun, Xie C, Si, H, Yang J (2017). CRISPR/Cas9-mediated genome editing in plants. Methods 121-122:94-102. |
|
|
Xiaonan M, Zhang X, Liu H, Li Z (2020). Highly efficient DNA-free plant genome editing using virally delivered CRISPR-Cas9. Nature Plants 6(7):773-779. |
|
|
Xingliang M, Mau M, Sharbel TF (2018). Genome Editing for Global Food Security. Trends in Biotechnology 36(2):123-127. |
|
|
Xingliang M, Zhu Q, Chen Y, Liu YG. (2016). CRISPR/Cas9 Platforms for Genome Editing in Plants: Developments and Applications. Molecular Plant 9(7):961-974. |
|
|
Macovei A, Sevilla NR, Cantos C, Jonson GB, Slamet-Loedin I, ?ermák T, Chadha-Mohanty P (2018). Novel alleles of rice eIF4G generated by CRISPR/Cas9-targeted mutagenesis confer resistance to Rice tungro spherical virus. Plant Biotechnology Journal 16(11):1918-1927. |
|
|
Makarova KS, Haft DH, Barrangou R, Brouns SJJ, Charpentier E, Horvath P, Koonin EV (2011). Evolution and classification of the CRISPR-Cas systems. Nature Reviews Microbiology 9(6):467-477. |
|
|
Makarova KS, Wolf YI, Alkhnbashi OS, Costa F, Shah SA, Saunders SJ, Koonin EV (2015). An updated evolutionary classification of CRISPR-Cas systems. Nature Reviews Microbiology 13(11):722-736. |
|
|
Malzahn A, Lowder L, Qi Y (2017). Plant genome editing with TALEN and CRISPR. Cell and Bioscience 7(1):1-18. |
|
|
Manghwar H, Lindsey K, Zhang X, Jin S (2019). CRISPR/Cas System: Recent Advances and Future Prospects for Genome Editing. Trends in Plant Science 24(12):1102-1125. |
|
|
Mao Y, Zhang H, Xu N, Zhang B, Gou F, Zhu JK (2013). Application of the CRISPR-Cas system for efficient genome engineering in plants. Molecular Plant 6(6):2008-2011. |
|
|
Martin AS, Salamango DJ, Serebrenik AA, Shaban NM, Brown WL, Harris RS (2019). A panel of eGFP reporters for single base editing by APOBEC-Cas9 editosome complexes. Scientific Reports 9(1):1-8. |
|
|
McFarlane GR, Whitelaw CBA, Lillico SG (2018). CRISPR-Based Gene Drives for Pest Control. Trends in Biotechnology 36(2):130-133. |
|
|
Mehta D, Stürchler A, Hirsch-Hoffmann M, Gruissem W, Vanderschuren H (2018). CRISPR-Cas9 interference in cassava linked to the evolution of editing-resistant geminiviruses. BioRxiv pp. 1-10. |
|
|
Mekler V, Kuznedelov K, Severinov K (2020). Quantification of the affinities of CRISPR-Cas9 nucleases for cognate protospacer adjacent motif (PAM) sequences. Journal of Biological Chemistry 295(19):6509-6517. |
|
|
Mikkelsen MD, Halkier BA (2003). Metabolic engineering of valine- and isoleucine-derived glucosinolates in arabidopsis expressing CYP79D2 from cassava. Plant Physiology 131(2):773-779. |
|
|
Molina R, Sofos N, Montoya G (2020). Structural basis of CRISPR-Cas Type III prokaryotic defence systems. Current Opinion in Structural Biology 65:119-129. |
|
|
Musunuru K (2017). The hope and hype of CRISPR-Cas9 genome editing: A review. JAMA Cardiology 2(8):914-919. |
|
|
Nakayama T, Fish MB, Fisher M, Oomen-Hajagos J, Thomsen GH, Grainger RM (2013). Simple and efficient CRISPR/Cas9-mediated targeted mutagenesis in Xenopus tropicalis. Genesis 51(12):835-843. |
|
|
Nambiar TS, Billon P, Diedenhofen G, Hayward SB, Taglialatela A, Cai K, Ciccia A (2019). Stimulation of CRISPR-mediated homology-directed repair by an engineered RAD18 variant. Nature Communications 10(1):1-13. |
|
|
Nassar NMA (2002). Cassava, Manihot esculenta Crantz, genetic resources: Origin of the crop, its evolution and relationships with wild relatives. Genetics and Molecular Research 1(4):298-305. |
|
|
Nocker S Van, Gardiner SE (2014). Breeding better cultivars, faster: Applications of new technologies for the rapid deployment of superior horticultural tree crops. Horticulture Research 1(1):1-8. |
|
|
O'Connor DJ, Wright GC, Dieters MJ, George DL, Hunter MN, Tatnell JR, Fleischfresser DB (2013). Development and Application of Speed Breeding Technologies in a Commercial Peanut Breeding Program. Peanut Science 40(2):107-114. |
|
|
Odipio J, Alicai T, Ingelbrecht I, Nusinow DA, Bart R, Taylor NJ (2017). Efficient CRISPR/cas9 genome editing of phytoene desaturase in cassava. Frontiers in Plant Science 8:1780. |
|
|
Odipio J, Alicai T, Nusinow D, Bart R, Taylor N (2018). CRISPR/Cas9-mediated Disruption of Multiple TFL1-like Floral Repressors Activates Flowering in Cassava. In In Vitro Cellular and Developmental Biology-Animal 54:S47-S47. 233 Spring St, New York, NY 10013 Usa: Springer. |
|
|
Okada A, Arndell T, Borisjuk N, Sharma N, Watson-Haigh NS, Tucker EJ, Whitford R (2019). CRISPR/Cas9-mediated knockout of Ms1 enables the rapid generation of male-sterile hexaploid wheat lines for use in hybrid seed production. Plant Biotechnology Journal 17(10):1905-1913. |
|
|
Palareti G, Legnani C, Cosmi B, Antonucci E, Erba N, Poli D, Tosetto A (2016). Comparison between different D-Dimer cutoff values to assess the individual risk of recurrent venous thromboembolism: Analysis of results obtained in the DULCIS study. International Journal of Laboratory Hematology 38(1):42-49. |
|
|
Pardo B, Gómez-González B, Aguilera A (2009). DNA double-strand break repair: How to fix a broken relationship. Cellular and Molecular Life Sciences 66(6):1039-1056. |
|
|
Piero Peisach E, Carl P, Robert G (2001). Design And Selection Ofnovel Cys2his2 Zinc Finger Proteins. Annual Review of Biochemistry 70(1):291-321. |
|
|
Peng R, Lin G, Li J (2016). Potential pitfalls of CRISPR/Cas9-mediated genome editing. FEBS Journal 283(7):1218-1231. |
|
|
MN (2013). Regeneration and RNAi-mediated downregulation of cyano-glycoside biosynthesis in cassava 167. |
|
|
Pinilla-Redondo R, Mayo-Muñoz D, Russel J, Garrett RA, Randau L, Sørensen SJ, Shah SA (2020). Type IV CRISPR-Cas systems are highly diverse and involved in competition between plasmids. Nucleic Acids Research 48(4):2000-2012. |
|
|
Prochnik S, Marri PR, Desany B, Rabinowicz PD, Kodira C, Mohiuddin M, Rounsley S (2012). The Cassava Genome: Current Progress, Future Directions. Tropical Plant Biology 5(1):88-94. |
|
|
Qin L, Li J, Wang Q, Xu Z, Sun L, Alariqi M, Jin S (2020). High-efficient and precise base editing of C•G to T•A in the allotetraploid cotton (Gossypium hirsutum) genome using a modified CRISPR/Cas9 system. Plant Biotechnology Journal 18(1):45-56. |
|
|
Ray DK, Mueller ND, West PC, Foley JA (2013). Yield Trends Are Insufficient to Double Global Crop Production by 2050. PLoS ONE 8(6):e66428. |
|
|
Razzaq A, Saleem F, Kanwal M, Mustafa G, Yousaf S, Arshad HMI, KhanJoyia FA (2019). Modern trends in plant genome editing: An inclusive review of the CRISPR/Cas9 Toolbox. International Journal of Molecular Sciences 20(16). |
|
|
Ricroch A (2019). Global developments of genome editing in agriculture. Transgenic Research 28(2):45-52. |
|
|
Rouillon C, Zhou M, Zhang J, Politis A, Beilsten-Edmands V, Cannone G, White MF (2013). Structure of the CRISPR interference complex CSM reveals key similarities with cascade. Molecular Cell 52(1):124-134. |
|
|
Sansbury BM, Hewes AM, Kmiec EB (2019). Understanding the diversity of genetic outcomes from CRISPR-Cas generated homology-directed repair. Communications Biology 2(1):1-10. |
|
|
Savary S, Willocquet L, Pethybridge SJ, Esker P, McRoberts N, Nelson A (2019). The global burden of pathogens and pests on major food crops. Nature Ecology and Evolution 3(3):430-439. |
|
|
Savic N, Ringnalda F, Bargsten K, Li Y, Berk C, Hall J, Schwank G (2017). Covalent linkage of the DNA repair template to the CRISPR/Cas9 complex enhances homology-directed repair. |
|
|
Schenke D, Cai D (2020). Applications of CRISPR/Cas to Improve Crop Disease Resistance: Beyond Inactivation of Susceptibility Factors. Iscience 23(9):101478. |
|
|
Schmidt C, Pacher M, Puchta H (2019). DNA break repair in plants and its application for genome engineering. Methods in Molecular Biology 1864:237-266. |
|
|
Sharma S, Kaur R, Singh A (2017). Recent advances in CRISPR/Cas mediated genome editing for crop improvement. Plant Biotechnology Reports 11(4):193-207. |
|
|
Shen H, Strunks GD, Klemann BJPM, Hooykaas PJJ, de Pater S (2017). CRISPR/Cas9-induced double-strand break repair in Arabidopsis nonhomologous end-joining mutants. G3: Genes, Genomes, Genetics 7(1):193-202. |
|
|
Shimada T (1978). Plant regeneration from the callus induced from wheat embryo. The Japanese Journal of Genetics 53(5):371-374. |
|
|
Shmakov S, Abudayyeh OO, Makarova KS, Wolf YI, Gootenberg JS, Semenova E, Koonin EV (2015). Discovery and Functional Characterization of Diverse Class 2 CRISPR-Cas Systems. Molecular Cell 60(3):385-397. |
|
|
Shmakov S, Smargon A, Scott D, Cox D, Pyzocha N, Yan W, Koonin EV (2017a). Diversity and evolution of class 2 CRISPR-Cas systems. Nature Reviews Microbiology 15(3):169-182. |
|
|
Shmakov S, Smargon A, Scott D, Cox D, Pyzocha N, Yan W, Koonin EV (2017b). Diversity and evolution of class 2 CRISPR-Cas systems. Nature Reviews Microbiology 15(3):169-182. |
|
|
Siritunga D, Sayre RT (2003). Generation of cyanogen-free transgenic cassava. Planta 217(3):367-373. |
|
|
Stinson BM, Moreno AT, Walter JC, Loparo JJ (2020). A Mechanism to Minimize Errors during Non-homologous End Joining. Molecular Cell 77(5):1080-1091. |
|
|
Symington LS, Gautier J (2011). Double-strand break end resection and repair pathway choice. Annual Review of Genetics 45:247-271. |
|
|
Tang XD, Gao F, Liu MJ, Fan QL, Chen DK, Ma WT (2019). Methods for enhancing clustered regularly interspaced short palindromic repeats/Cas9-mediated homology-directed repair efficiency. Frontiers in Genetics 10:551. |
|
|
Tashkandi M, Ali Z, Aljedaani F, Shami A, Mahfouz MM (2018). Engineering resistance against Tomato yellow leaf curl virus via the CRISPR/Cas9 system in tomato. Plant Signaling and Behavior 13(10):1-7. |
|
|
Tomlinson KR, Bailey AM, Alicai T, Seal S, Foster GD (2018). Cassava brown streak disease: historical timeline, current knowledge and future prospects. Molecular Plant Pathology 19(5):1282-1294. |
|
|
Tsai SQ, Zheng Z, Nguyen NT, Liebers M, Topkar VV, Thapar V, Joung JK (2015). GUIDE-seq enables genome-wide profiling of off-target cleavage by CRISPR-Cas nucleases. Nature Biotechnology 33(2):187-198. |
|
|
Tyagi S, Kumar R, Kumar V, Won SY, Shukla P (2021). Engineering disease resistant plants through CRISPR-Cas9 technology. GM Crops and Food 12(1):125-144. |
|
|
Ueta R, Abe C, Watanabe T, Sugano SS, Ishihara R, Ezura H, Osakabe K (2017). Rapid breeding of parthenocarpic tomato plants using CRISPR/Cas9. Scientific Reports 7(1):1-8. |
|
|
Veley KM, Okwuonu I, Jensen G, Yoder M, Taylor NJ, Meyers BC, Bart RS (2021). Gene tagging via CRISPR-mediated homology-directed repair in cassava. G3 Genes|Genomes|Genetics 11(4)::jkab028. |
|
|
Walsh RM, Hochedlinger K (2013). A variant CRISPR-Cas9 system adds versatility to genome engineering. Proceedings of the National Academy of Sciences of the United States of America 110(39):15514-15515. |
|
|
Wang M, Wang S, Liang Z, Shi W, Gao C, Xia G (2018). From Genetic Stock to Genome Editing: Gene Exploitation in Wheat. Trends in Biotechnology 36(2):160-72. |
|
|
Wang W, Pan Q, He F, Akhunova A, Chao S, Trick H, Akhunov E (2018). Transgenerational CRISPR-Cas9 Activity Facilitates Multiplex Gene Editing in Allopolyploid Wheat. The CRISPR Journal 1(1):65-74. |
|
|
Watters K (2018). The CRISPR Revolution: Potential Impacts on Global Health Security. |
|
|
Williams BO, Warman ML (2017). CRISPR/CAS9 Technologies. Journal of Bone and Mineral Research 32(5):883-888. |
|
|
Chhotaray C, Tan Y, Mugweru J, Islam MM, Hameed HA, Wang S, Lu Z, Wang C, Li X, Tan S, Liu J (2018). Advances in the development of molecular genetic tools for Mycobacterium tuberculosis. Journal of Genetics and Genomics 45(6):281-297. |
|
|
Ye L, Wang C, Hong L, Sun N, Chen D, Chen S, Han F (2018). Programmable DNA repair with CRISPRa/i enhanced homology-directed repair efficiency with a single Cas9. Cell Discovery 4(1):1-12. |
|
|
Yin K, Han T, Liu G, Chen T, Wang Y, Yu AYL, Liu Y (2015). A geminivirus-based guide RNA delivery system for CRISPR/Cas9 mediated plant genome editing. Scientific Reports 5:1-10. |
|
|
Young K, Gasior SL, Jones S, Wang L, Navarro P, Vickroy B, Barrangou R (2019). The repurposing of type I-E CRISPR-Cascade for gene activation in plants. Communications Biology 2(1):1-7. |
|
|
Zaidi SSeA, Mukhtar MS, Mansoor S (2018). Genome Editing: Targeting Susceptibility Genes for Plant Disease Resistance. Trends in Biotechnology 36(9):898-906. |
|
|
Zhang Q, Xing HL, Wang ZP, Zhang HY, Yang F, Wang XC, Chen QJ (2018). Potential high-frequency off-target mutagenesis induced by CRISPR/Cas9 in Arabidopsis and its prevention. Plant Molecular Biology 96(4-5):445-456. |
|
|
Zhang ZT, Jiménez-Bonilla P, Seo SO, Lu T, Jin YS, Blaschek HP, Wang Y (2018). Bacterial genome editing with CRISPR-Cas9: Taking clostridium beijerinckii as an example. Methods in Molecular Biology 1772:297-325. |
|
|
Zhou H, Liu B, Weeks DP, Spalding MH, Yang B (2014). Large chromosomal deletions and heritable small genetic changes induced by CRISPR/Cas9 in rice. Nucleic Acids Research 42(17):10903-10914. |
|
|
ZHOU W, MA Q, ZHANG P, WANG L, WANG H, XU J, YANG J (2013). Key Scientific Questions and Recent Advances in Cassava Molecular Breeding. Scientia Sinica Vitae 43(12):1082-1089. |
|
|
Zuo Z, Zolekar A, Babu K, Lin VJ, Hayatshahi HS, Rajan R, Liu J (2019). Structural and functional insights into the bona fide catalytic state of Streptococcus pyogenes Cas9 HNH nuclease domain. Elife 8:e46500. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


