Full Length Research Paper

ABSTRACT
INTRODUCTION
The Arabian Sand Gazelles (Gazella subgutturosa marica) is classified as vulnerable (VU) on the IUCN Red List (IUCN Red, 2009) and listed under appendix II of the convention on migratory species (CMS, 2005). Scientific references on the Arabian Sand Gazelles concern only the ethological aspects and reintroduction as well as veterinary issues and diseases (KKWRC, 2008). Yet, the information on reproductive physiology is still lacking. A few reports concern these issues (Al-Eissa and Alhomida, 1997; Al-Eissa, 2007a, 2007b) as the knowledge on reproductive male characteristics is crucial for protection of the particular species from extinction.
Testosterone is a steroid hormone from the androgen group and is found in mammals, reptiles, birds, and other vertebrates. In mammals, testosterone is secreted primarily by the testicles of males. It is the principal male sex hormone and an anabolic steroid (Alder and Cox, 1983). Follicle-stimulating hormone (FSH) is a glycoprotein gonadotropin secreted by the anterior pituitary in response to gonadotropin-releasing hormone (GnRH), which is released by the hypothalamus. The same pituitary cell also secretes luteinizing hormone (LH), another gonadotropin. FSH and LH are composed of alpha and beta subunits. The specific beta subunit confers the unique biologic activity. FSH and LH bind to receptors in the testis and ovary and regulate gonadal function by promoting sex steroid production and gametogenesis (Grover and Malm, 2005; Bassil and Morley, 2009). FSH stimulates testicular growth and enhances the production of an androgen-binding protein by the Sertoli cells, which are a component of the testicular tubule necessary for sustaining the maturing sperm cell. Hence, maturation of spermatozoa requires FSH and LH. Stimulation of LH estrogen and progesterone production from the ovary.
FSH and LH secretion are affected by a negative feedback from sex steroids. Inhibin also has a negative feedback on FSH selectively. High-dose testosterone or estrogen therapy suppresses FSH and LH. Primary gonadal failure in men and women leads to high levels of FSH and LH, except in selective destruction of testicular tubules with subsequent elevation of only FSH, as in Sertoli-cell-only syndrome. Previous study indicated that the whole reproductive system of fallow deer is directly controlled by the photoperiod and that related morpho-logical and physiological processes are indirectly determined by photoperiodic changes (Rolef and Fisher, 1990).
Seasonal levels of LH, FSH, testosterone (T) and prolactin (PRL) were determined in plasma of adult male Southern Pudu deer (Pudu puda) kept in Concepcion, FSH concentrations remained at peak levels (54 to 63 ng/ml) from December to March; minimal values (25 to 33 ng/ml) were detected from April until October. Testosterone levels exhibited two, almost equal peaks; the first peak (2.8 ng/ml) was detected in March and the second one (2.7 ng/ml) in October. Both T peaks were preceded by an earlier elevation of LH in February and July (both around 1.3 ng/ml). During the fall, only the alpha male exhibited a sharp peak of T (8.4 ng/ml), whereas in the spring five out of six bucks demonstrated an increase of T levels. Two peaks of LH and T and the 4 months of elevated FSH may be related to a long period of spermatogenesis observed in this species (George et al., 1996). The role of the adrenocortical system in the regulation of plasma levels of reproductive hormones was investigated and plasma levels of LH, testosterone (T), FSH, prolactin (PRL) and androstenedione were determined in male white-tailed deer after ACTH, The result showed no effect on FSH and PRL levels (Bubenik et al., 1990).
MATERIALS AND METHODS
All of the experimental procedures were conducted in King Khalid Wildlife Research Center (KKWRC) (25°03’N, 46°45’E), Saudi Wildlife Commission (SWC), Riyadh, Saudi Arabia.
Animals
Ten (10) males of Arabian Gazelles G. subgutrosa marica aged 2 to 4 years (average 15 to 18 kg live weight each) served as serum donors. The males were kept in enclosure 100 x 100 m and all were good health well feed mainly on dry alfalfa, some concentrated pellets and water.
Radioimmunoassay (RIA) for hormones
Testosterone, concentrations in the serum was quantified by commercial available RIA kits (Immunotech, France). The levels of FSH, LH were measured by rate specific FSH and LH RIA kits (DRG international, Germany).
Semen collection and processing
Semen collection
Semen was collected from the males by using an electroejaculation procedure described by Al-Eissa (2007) and Al-Eissa et al. (2007; 2009) (Figure 1). The animals were anaesthetized using Xylazine (8.4±1.5 mg kg-1 b.wt.) and ketamine hydrochloride 6.9±1.5 mg kg-1 b.wt.). During the electroejaculation procedure, which typically involved <10 stimulations up to maxima of 4.5 to 7.0 volts, also similar method was described by Holt et al. (1988; 1996). Semen analysis was carried out immediately after collection. Seminal plasma sodium (Na), potassium (K) and chlorides (CL), were assessed with commercial kits (Labtest Diagnostica, Brazil) using an electrolyte analyzer (Iselab Eletrolyte Analyser, Brazil) to determine seminal plasma concentrations of each element. Qualitative measurement of fructose in seminal fluid was carried out by Resorcinol method (Fauser et al., 1990).
Statistical analysis
All data were expressed as mean±SD. The difference in serum concentration of FSH, LH, and Testosterone was evaluated by one-way ANOVA. Differences were considered to be significant at p<0.05.
RESULTS
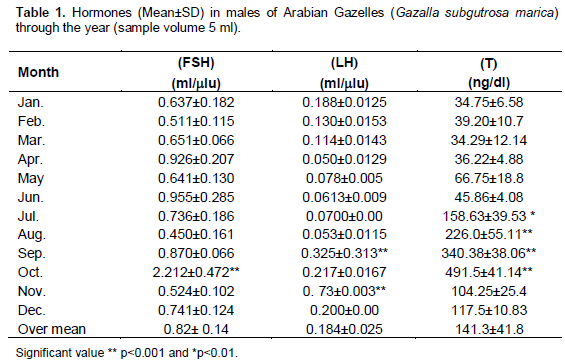
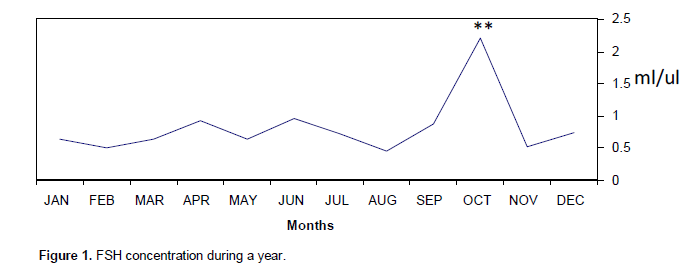
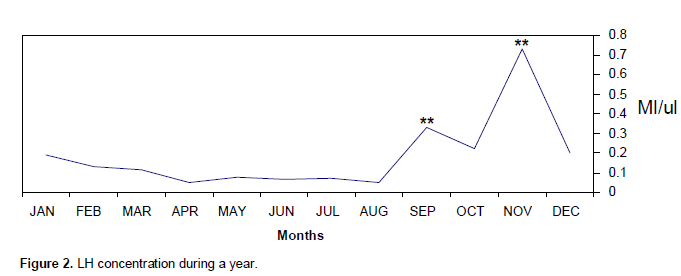
The concentrations of serum levels of Testosterone T, FSH and LH were measured by Radioimmunoassay RIA method. The obtained results fluctuated throughout the year with high levels during mating season as shown in Table 1. The mean of FSH hormone was significantly (p<0.001) increased compared with other months and reached its peak (2.212 mµ) in October which is the sexual season Figure 1. Also, LH hormone increased one month before FSH and significantly (p<0.001) increased compared with other months and reached its peak 0.73 mµ during September and November Figure 2. In contrast, testosterone hormones significantly (p<0.01) increased from May until August and significantly (p<0.001) increased to reached its peak in September (340.38 ng/dl) and October (491.5 ng/dl), respectively, later on testosterone quick dropped in November (Table 1 and Figure 3).
Plasma parameters
Some inorganic parameters of semen plasma were detected. These parameters of five males of Arabian Gazelles G. subgutrosa marica are illustrated in Table 2. The concentrations of sodium, potassium, chloride ele-ments and fructose compound were measured during four months; February, May, August and November. Semen was collected every month from five males. Mean concentration of Sodium was 141±3.5 mM/L and the highest concentration was 147 mM/L in November. Mean concentration of calcium was 7.2±0.15 mM/L and the highest concentration was 7.2±0.15 mM/L in November. Mean concentration of chloride was 141±3.5 mM/L and the highest concentration was 129 mM/L in November. Mean concentration of fructose was 652.75±29 mg/dL and the highest concentration was 680 mg/dL in November. No significant changes in Semen plasma parameters were observed (Table 2).

Semen
Table 1 shows hormones (mean±SD) in males of Arabian Gazelles (G. subgutrosa marica) through the year (sample volume 5 ml). Table 2 shows semen plasma parameters (mean±SD) in males of Arabian Gazelles (G. subgutrosa marica) during the year (sample volume 0.5 ml).
DISCUSSION
The Arabian Sand Gazelles (G. subgutturosa marica) is classified as vulnerable (VU) on the IUCN Red List (IUCN Red, 2009). Scientific references on the Arabian Sand Gazelles concern only the ethological aspects and reintroduction as well as veterinary issues and diseases (KKWRC, 2008). The limitation of this study was the scant number of valued subjects (Chemineau, 1986). The information on reproductive physiology is still lacking. A few reports concerned these issues (Al-Eissa and Alhomida, 1997; Al-Eissa ,2007a; Al-Eissa et al., 2007b). Some studies described the reproductive season of Arabian Sand Gazelles and compared its reproductive cycles to Persian gazelle (G. subgutturosa subgutturosa) (Sempere, 2001). In the present study, concentration of FSH, LH and T slightly increased pre-sexual period. As indicated in result, FSH hormones increased rapidly in October during mating season as shown in Table 1, and this revealed the important role of sex hormones in sexual period and also, its role in spermatogenesis. This data go in line with previous study on Pudu puda that showed increment of FSH during sexual period (George et al., 1996).
In contrast, LH hormone increased one month before FSH; this means that LH activated testosterone to enhance sperm premature. Again, this result agrees with that of George et al. (1996) on (pudu puda). Basically, increasing of Testosterone during three months of sexual period reflected on its fundamental functions that related to formation and preserve of spermatogenesis. This result supported (Gundogan, 2005) study that conducted on the sheep. There are evidences that a season influenced on both semen production and characteristics in some antelope breeds (Asher et al., 1993). In other antelope like goitred gazelles (G. subgutturosa subgutturosa) living in an arid environment conditions, intense sexual activity was observed during the breeding season (November to December) (Mambetdjumaev, 1970; Gorelov, 1972; Djevnerov, 1984; Blank, 1985).
Previous study reported morphometric, plasma testosterone, LH concentrations and seminal parameters in European fallow deer (Dama dama dama) and other genotypes. These results exhibited significant displacement between genotypes among year and revealed markedly seasonal patterns of secretion of testosterone and LH, with hybrid males exhibiting an apparent earlier onset of high-amplitude testosterone in February compared to those occurring in April for European males. Taken together, the data indicate strongly that the Mesopotamian influence is evident in the earlier attainment of sexual development and fertility in late summer and autumn, and earlier onset of sexual quiescence in spring (Asher et al, 1996).
The obtained data illustrated that the concentration of some inorganic semen compounds are non-significant increased during sexual period compared with other months. This result agrees with other studies that revealed the role of semen plasma to protect and enhance motility and activity of sperms (Gundogan, 2005) and protect sperms from cold shock (Barrios et al., 2000). However, it is very important to protect endangers species by all means of reproductive biotechnologies and understand the reproductive physiology of these animals, in order to increase the number of these threaten animals. In addition, extend the captive population and establish the more cryopreservation of genetic resources banks. Al-Eissa et al. (2007a) reported a breeding season in late summer and autumn with high quantity of semen volume and progressive spermatozoa. In gazelles males the breeding season in temperate areas seems to be different from the mountain gazelle. A possible hypothesis could be related to the Mountain gazelle different sensitivity to photoperiod or other influences on melatonin release (Al-Eissa, 2007a). Semen characteristics of mountain gazelle were highly influenced by season. The acceptable semen production to try cryopreservation programs was produced in October to December, but more investigations are needed in order to identify the exact breeding period. Frozen semen could be utilized in artificial insemination, for genetic studies, to increase genotypic variability and resolve the problem of rupicaprine species extinction (Al-Eissa, 2007a).
CONCLUSION
In conclusion, sexual hormones were strongly influenced by season. The results from this study suggest more investigation on the reproductive physiological research in order to detect the suitable period for semen collection of Arabian Sand Gazelles (G. subgutturosa marica) in Saudi Arabia. Also, to detect the suitable period for breeding in order to protect this animals and establish Cryopreservation of genetic resources banks in in Saudi Arabia for manipulation and long-term preservation.
CONFLICT OF INTERESTS
The author(s) have not declared any conflict of interests.
ACKNOWLEDGEMENT
The author extends his appreciation to the Deanship of Scientific Research at King Saud University for funding the work through the research group project No. RGP-018.
REFERENCES
|
Alder E and Cox J (1983). Breast feeding and post-natal depression. Psychosom. Res. 27(2):139-144. Crossref |
||||
| Al-Eissa MS (2007a). Assessment of reproductive efficiency of the Arabian Sand Gazelle males (Gazella subgutturosa marica) and semen cryopreservation". PhD Thesis, King Saud University, College of Science, Department of Zoology. Al-Eissa MS, Alhamidi AR, Khandil S (2007b). Semen cryopreservation using triladyl and tris diluents of the Arabian Sand Gazelle males (Gazelle Subgutturosa marica)". Arabian Gulf J. Sci. Res. (4) 25: 199-206. | ||||
|
Al-Eissa MS, Alhomida AS (1997). A study of the distribution of total, free, short-chain acyl and long-chain acyl carnitine in whole blood and plasma of Arabian Sand Gazelle (Gazelle Subgutturosa marica"). Comp. Haematol. Int. 1:65-69. Crossref |
||||
|
Asher GW, Berg DK, Beaumont S, Morrow CJ, O'Neill KT and Fisher MW (1996). Comparison of seasonal changes in reproductive parameters of adult male European fallow deer (Dama dama dama) and hybrid Mesopotamian x European fallow deer (D. d. mesopotamica x D. d. dama). Anim. Reprod. Sci. 45(3):201-215. Crossref |
||||
|
Asher GW, Fisher MW, Fennessy PF, Mackintosh CG, Jabbour HN, Morrow CJ (1993). Oestrous synchronization, semen collection and artificial insemination of farmed red deer (Cervus elaphus) and fallow deer (Dama dama). Anim. Reprod. Sci. 33: 241-265. Crossref |
||||
|
Barrios B, Perez R, Gallego M, Tato A, Muino-Blanco. (2000). Seminal plasma proteins revert the cold-shock damage on ram sperm membrane. Biol. Reprod. 63:1531-1537. Crossref |
||||
| Bassil N, Morley J (2009). Endocrine aspects of healthy brain aging. Clin. Geriatr. Med. 1:57-74. | ||||
| Bubenik G, Smith J, Pomerantz D, Schams D (1990). Plasma LH, FSH, testosterone, prolactin and androstenedione in male white-tailed deer after ACTH and dexamethasone administration. Comp. Biochem. Physiol. A Comp. Physiol. 95(1):163-169. | ||||
|
Chemineau P (1986). Sexual behaviour and gonadal activity during the year in the tropical Creole meat goat. II. Male mating behaviour, testis diameter, ejaculate characteristics and fertility. Reprod. Nutr. Dev. 26: 453-460. Crossref |
||||
|
CMS (2005). Convention on Migratory Species. |
||||
| Djevnerov VV (1984). Goitred Gazelle of Barsakelmes Island. Alma-Ata: Nauka. p. 144. | ||||
|
Fauser BC, Bogers JW, Hop WC, De-Jong FH (1990). Bioactive and immunoreactive FSH in serum of normal and oligospermic men. Clin. Endocrinol. Oxf. 32:433-442. Crossref |
||||
| George A, Eugenia R, Dieter S, Alejandro L, Luded B (1996). Seasonal Levels of LH, FSH, testosterone and prolactin in male pudu (Pudu puda) . Comp. Biochem. Physiol. B Biochem. Mol. Biol. 115B (4): 417-420. | ||||
| Grover G, Malm J (2005). Selective thyroid hormone agonists: A strategy for treating metabolic syndrome. Metab. Syndr. 2 (2): 137-142. | ||||
| IUCN Red List (2009). View | ||||
| KKWRC (2008). National Commission for wildlife Conservation and Development (NCWCD). Riyadh, Saudi Arabia. | ||||
| Mambetdjumaev A (1970). Goitred Gazelles. Tashkent, Fan. p.196. | ||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


