ABSTRACT
The combined action of both endogenous and exogenous factors stimulates the production of free radicals and reactive oxygen species (ROS) in cells. They are neutralized by an elaborate antioxidant defense system, superoxide dismutase (SOD) being the first line of enzymes involved in ROS scavenging. However, when ROS production cannot be counteracted by cellular antioxidant mechanisms oxidative stress happens, inducing damages in proteins, lipids and DNA. It had been suggested that antioxidant supplementation may help to reduce oxidative stress. TetraSOD® is a unique marine healthy and functional ingredient that is produced under patent-protected technology, which exhibits a balanced nutritional composition and a significantly high SOD activity (>30,000 U/g). In this work, the antioxidant bioactivity of TetraSOD® in muscle cells (Normal Human Skeletal Muscle Myoblasts, HSMMs) was examined using an in vitro approach. Three different doses of SOD activity in TetraSOD® extracts were tested in cells: 30, 150 and 1500 U/ml. A statistically significant induction of the primary antioxidant enzyme activities, SOD, glutathione peroxidase (GPx), and catalase (CAT) was observed at the two higher doses in relation to the control. These results were further related to the expression of a selected set of genes involved in response against oxidative stress. A significant up-regulation of SOD1, SOD2, GPx1, CAT, NRF2 and HMOX1 was detected after TetraSOD® treatment with regard to the control, exhibiting different patterns in response to the applied dose to cells. Overall, the results obtained in this study might represent an important contribution to the understanding of the molecular basis underlying the potential cytoprotective effects induced by TetraSOD® consumption.
Key words: TetraSOD®, Tetraselmis chuii, oxidative stress, antioxidant, gene expression, enzyme activity, SOD, NRF2.
Cells produce reactive oxygen species (ROS) as a consequence of the own metabolic processes, the superoxide anion being the precursor of all other ROS (Romao, 2015). The three main in vivo sources for superoxide anion are the mitochondrial respiratory chain complexes, the nicotinamide adenine dinucleotide phosphate-oxidase, and xanthine oxidases. ROS can provoke damages in proteins, lipids and DNA when
antioxidant capabilities are overwhelmed by the burden of ROS, a state known as “oxidative stress” (Tebay et al., 2015). To detoxify ROS cells have developed an elaborate antioxidant defense system in which superoxide dismutase (SOD) is the first line of enzymes involved in scavenging of ROS. This enzyme reduces superoxide anion to hydrogen peroxide (H2O2), and then glutathione peroxidase (GPx) and catalase (CAT) convert H2O2 into H2O and O2 (Ighodaro and Akinloye, 2018).
Occurrence of oxidative stress as a consequence of both endogenous metabolism and exogenous factors (diet, pollution, UV radiation, smoking, mental stress…) is considered to be involved in a number of diseases such as cancer, atherosclerosis, diabetes, chronic inflammation, and neurodegenerative and cognitive disorders (Fiedor and Burda, 2014). In this scenario, dietary factors seem to play a pivotal role in the regulation of the oxidant status as a diet low in antioxidants contributes to oxidative stress (Décordé et al., 2010). A wide range of studies have examined the beneficial effects of antioxidant supplementation to reduce oxidative stress; particularly, SOD is probably one of the antioxidant proteins most extensively addressed (Carillon et al., 2013a; Romao, 2015; Stephenie et al., 2020). In this regard, an increase in endogenous antioxidant enzymes (not only SOD but also GPx and CAT) has been observed after SOD supplementation both at protein and activity levels, supporting the hypothesis that exogenous SOD could induce endogenous antioxidant enzymes (Vouldoukis et al., 2004; Nelson et al., 2006; Carillon et al., 2013a, b, 2014; Romao, 2015). However, the molecular mechanisms underlying such induction processes have not been elucidated yet.
TetraSOD® is a unique marine healthy and functional ingredient that is grown under patent-protected technology (Unamunzaga and Mantecón, 2015). In addition to its exhibited high SOD activity (>30,000 U/g), it shows a balanced nutritional composition containing essential fatty acids, vitamins, minerals, amino acids and pigments. With the aim to understand the physiological bases of the protective health effects of TetraSOD®, in vitro testing of bioactivity was assessed in human cells as a first step in the elucidation of mechanisms explaining such protection. Thus, the effect of TetraSOD® in the three main antioxidant enzyme activities (SOD, GPx and CAT) in Normal Human Skeletal Muscle Myoblasts (HSMMs) was determined. Moreover, to gain additional insights on the TetraSOD® effect at molecular level, the expression profile of a selected set of genes encoding for proteins involved in protection against oxidative stress was analyzed including: (i) the three distinct identified isoforms of SOD in mammals. Two of the isoforms have Cu and Zn in the catalytic center and exhibit different localization, being intracellular (Cu,Zn-SOD or SOD1) or extracellular (EC-SOD or SOD3). The last isoform has Mn as a cofactor (Mn-SOD or SOD2) and is localized to mitochondria (Zelko et al., 2002; Sheng et al., 2014); (ii) glutathione peroxidases, which couple oxidation of glutathione with detoxification of H2O2. The family comprises up to eight isoforms in mammals. Among them, both GPx1 and GPx4 are ubiquitous as have been identified in the cytosol, nucleus and mitochondria of the cells, and also exhibit a wide tissue distribution (Margis et al., 2008; Espinosa-Diez et al., 2015); (iii) the human catalase, which is an heme-containing peroxidase that forms a tetramer composed of four subunits, each one divided in four domains, and plays a predominant role in controlling the concentration of H2O2 (Putnam et al., 2000; Goyal and Basak, 2010); (iv) the heme oxygenase-1 (HMOX1) inducible phase II detoxifying enzyme, which is considered an important component involved in the adaptive and protective response to multiple oxidative insults (Solano and Arck, 2015; Liao et al., 2018); (v) the nuclear factor erythroid 2-related factor 2 (NRF2), which is a transcription factor that under physiological conditions is sequestered by binding to Kelch-like ECH associated protein (KEAP1), thus inhibiting translocation of NRF2 into the nucleus. When KEAP1 changes its conformation due to interaction with different inducers, NRF2 is released and translocated to the nucleus, regulating the cytoprotective response to oxidative stress through transcriptional activation of phase II detoxifying and antioxidant enzymes (Dinkova-Kostova and Talalay, 2008; Tkachev et al., 2011). Main results of this research are presented here in after.
Microalgae biomass (TetraSOD®)
TetraSOD® was produced by Fitoplancton Marino, S.L. using a patent protected technology. It corresponds to a lyophilized powder of the microalgae Tetraselmis chuii (T. chuii) strain CCFM 03 (belonging to the culture collection of Fitoplancton Marino, S.L.), which is grown in controlled outdoor close photobioreactors (proprietary process) under photoautotrophic conditions ensuring high SOD activity (>30,000 U/g) (patent protected).
Extract preparation and SOD activity determination in TetraSOD®
Extract was prepared from TetraSOD® using a proprietary methodology. A 10% dry weight solution in phosphate buffer (KH2PO4 200 mM pH=7.8) was disrupted using a high pressure homogenizer. Cell debris was removed by centrifugation at 20,000 g for 15 min at 4ºC, and the supernatant was recovered and stored at -80ºC until freeze-drying. An aliquot was first removed to determine SOD activity, which was quantified using the SOD Assay Kit-WST (Dojindo) according to the Technical Manual. The inhibition rate was determined by a kinetic method using a time range between 2 and 10 min, in which a very high linearity was observed (R2>0.99).
Cell culture and incubation with TetraSOD® extract
Normal human skeletal muscle myoblasts (HSMMs, Clonetics™) were purchased from Lonza (Walkersville, USA) and cultured according to instructions provided by the manufacturer. They were grown in SkGM™-2 BulletKit™ medium in a humidified incubator at 37°C with a 5% CO2 atmosphere. Cells were subcultured when they were 50 to 70% confluent and contained many mitotic figures throughout the flask.
Cells were incubated in 6-well plates for 24 h with TetraSOD® extract prepared at different dilutions (2 ml total volume per well) containing 30, 150 and 1500 U/ml of culture media. As a control treatment, phosphate buffer was employed. Afterwards, cells were rinsed twice with PBS and directly processed for enzyme activities determination or RNA isolation as described subsequently. The experiment was repeated twice, and in each one, treatment was performed in triplicate.
SOD, GPx and CAT activity determination in HSMMs cells
Cells were detached by gentle treatment with Trypsin/EDTA solution provided with the ReagentPack™ Subculture Reagents (Clonetics™). After cells were released, trypsin was neutralized with Trypsin Neutralizing Solution (Clonetics™). Thereafter, cells were washed twice in ice cold phosphate buffer by centrifugation at 220 g for 5 min at 4ºC. Then cells were resuspended in phosphate buffer containing 1% peroxide-free Triton X-100. Complete lysis was achieved by softly pipetting up and down several times. Finally, cell lysate was centrifuged at 20,000 g for 10 min at 4ºC, and the supernatant was recovered and immediately stored at -80ºC until use.
SOD activity was quantified using the SOD Assay Kit-WST (Dojindo) according to the Technical Manual as previously indicated. GPx activity (nmol/min/ml) was quantified using the Glutathione peroxidase activity kit (Enzo) as described in the manual provided by the manufacturer. CAT activity (nmol/min/ml) was determined using the Catalase Assay Kit (Cayman Chemical) following the kit booklet. In all instances, activity values were transformed to percentages in relation to the Control (without TetraSOD® incubation).
RNA isolation and gene expression analysis
Total RNA was isolated from HSMMs cells using the NucleoSpin® RNA Kit (Macherey-Nagel) in accordance with the manufacturer's protocols. A double on column DNase I treatment for 30 min was performed to avoid amplification of genomic DNA. Quality of RNA samples was first checked in agarose gels, and then A260/A280 and A260/A230 ratios (>2.1) were determined with a NanoDrop 2000 spectrophotometer (Thermo Scientific). Total RNA (1 μg) from each sample was reverse-transcribed with the iScript™ cDNA Synthesis kit (Bio-Rad) following the manufacturer's protocol. Lack of genomic DNA contamination was confirmed by PCR amplification of RNA samples in the absence of cDNA synthesis.
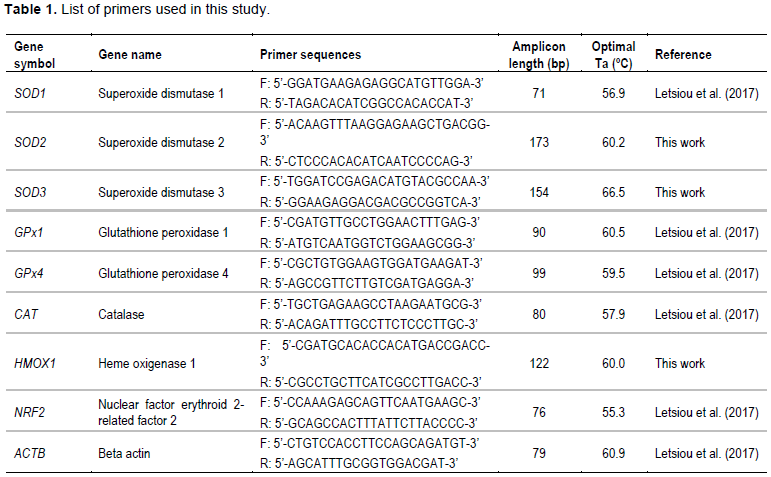
Real-time analysis was carried out on a CFX96™ Real-Time System (Bio-Rad) using specific primers for SOD1, SOD2, SOD3, GPx1, GPx4, CAT, HMOX1 and NRF2 (Table 1). Real-time reactions were accomplished in a 10 μL volume containing cDNA generated from 10 ng of original RNA template, 300 nM each of specific forward and reverse primers, and 5 μl of iQ™ SYBR® Green Supermix (Bio-Rad). The amplification protocol included an initial 7 min denaturation and enzyme activation step at 95°C, followed by 40 cycles of 95°C for 15 s and 60°C for 30 s. Each PCR assay was done in duplicate. For normalization of cDNA loading, all samples were run in parallel with the reference gene beta actin (ACTB) as previously reported (Letsiou et al., 2017). Relative mRNA expression was determined using the 2−(ΔΔCt) method (Livak and Schmittgen, 2001).
Statistical analysis
Results were expressed as mean ± SD. Significant differences were determined with the non-parametric Kruskal-Wallis test followed by Dunn's multiple comparisons test. These analyses were performed using the Prism v6 software (GraphPad Software). Significance was accepted at p<0.05.
Antioxidant enzyme activities after TetraSOD® treatment
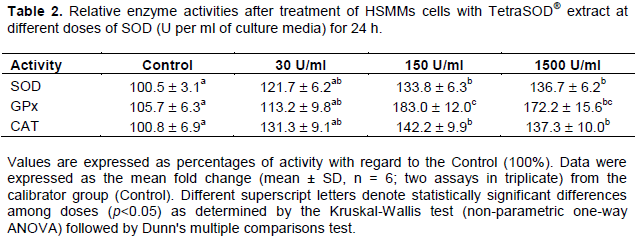
The effect of TetraSOD® on the three antioxidant enzyme activities SOD, GPx and CAT was analyzed. In all instances, the lowest dose of TetraSOD® did not provoke a statistically significant change in activity when compared to the Control. However, in both 150 and 1500 U/ml treatments a significant increase was detected in the three activities. In SOD, the increase ranged around 34% in 150 U/ml and 37% in 1500 U/ml in relation to the control. In GPx activity, the highest variation was observed, ranging between 83 and 72% at 150 and 1500 U/ml, respectively. And in CAT, the increase in activity values ranged between 42 and 37% at 150 and 1500 U/ml, respectively (Table 2).
TetraSOD® induces up-regulation of genes involved in antioxidant response
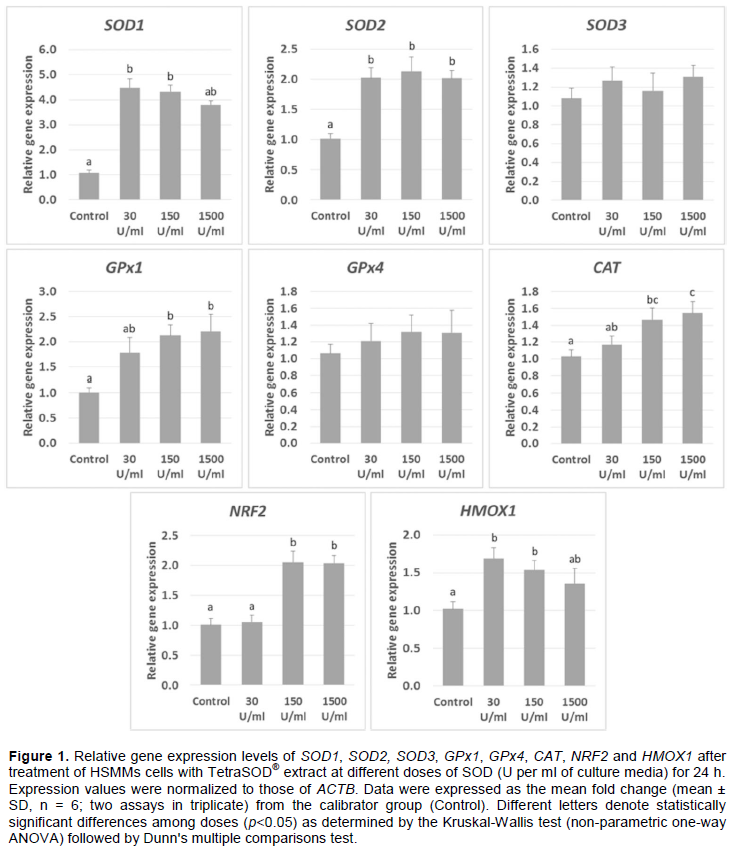
Incubation of HSMMs with TetraSOD® extract up-regulated some of the most relevant genes involved in cellular antioxidant response (Figure 1). With 30 U/ml, a significant 4.49-fold increase in SOD1 mRNAs with regard to the control was observed. At 150 U/ml it increased to 4.32-fold, and it was 3.80-fold higher (although not significant with regard to the control) at 1500 U/ml. No significant differences were found between any of the TetraSOD® treatments. Incubation of HSMMs cells with TetraSOD® also up-regulated SOD2 at the three concentrations tested, with an increase in transcript amounts ranging between 2.02-fold at 1500 U/ml and 2.14-fold at 150 U/ml, with no statistically significant differences between treatments. In contrast to SOD1 and SOD2, SOD3 did not exhibit differences in mRNA abundance between any of the treatments and the control.
Moreover, TetraSOD® induced a significant up-regulation of GPx1 at both 150 U/ml (2.14-fold higher than Control) and 1500 U/ml (2.23-fold increase). At 30 U/ml, a 1.85-fold increase was measured, although it was not statistically significant in relation to the control. In contrast, no significant changes were detected in GPx4 between any of the treatments and the control.
In addition, treatment of HSMMs cells with both 150 and 1500 U/ml of TetraSOD® provoked a significant increase in mRNA levels of CAT with regard to the control (1.47-fold and 1.55-fold, respectively). A non-statistically significant 1.17-fold increase in transcripts was observed at 30 U/ml, which was significant when compared to 1500 U/ml treatment.
Finally, when HMMS cells were incubated with TetraSOD® at 30 U/ml, no significant changes in NRF2 transcripts were detected. In contrast, a significant increase in mRNAs was measured both at 150 (2.05-fold) and 1500 (2.04-fold) U/ml. Moreover, a significant up-regulation of HMOX1 was observed after treatment with 30 and 150 U/ml, exhibiting 1.19- and 1.54-fold higher transcript amounts than the Control, respectively. However, no significant differences were observed at the highest TetraSOD® dose.
Oxidative stress occurs when molecular defense systems are not able to counteract oxidation caused by both endogenous processes (such as mitochondrial metabolism) and/or exogenous factors including chemicals, pollution or diet. Hence, it reflects an imbalance of any biological system to detoxify ROS or repair resulting damages in cell components (Tebay et al., 2015). In the present study, the antioxidant effect of TetraSOD® using an in vitro approach in HSMMs cells was investigated, and the underlying molecular mechanisms of antioxidant protection were further explored. In this regard, a significant induction in the activity of the antioxidant enzymes SOD, GPx and CAT was observed after TetraSOD® treatment. On this matter, a literature search revealed a wide range of compounds of different nature (peptides, chemical reagents, natural extracts…) able to induce activity of antioxidant enzymes in different cell systems. For instance, pretreatment of hippocampus cells with the cocaine- and amphetamine-regulated transcript (CART) peptide prevented the oxidative damage induced by amyloid-beta microinjection through a mechanism involving up-regulation of SOD activity (Jiao et al., 2018). An increase in not only SOD but also GPx activity has been observed in response to oxidative damage induced by UV-A on normal human dermal fibroblasts after treatment with polypeptide from Chlamys farreri (Han et al., 2004). SOD activity was also up-regulated in human skin fibroblasts treated with the flavonoid baicalin in response to UV-A radiation (Zhou et al., 2012). Similar effects have been reported regarding SOD and CAT activity in human umbilical vein endothelial cells pretreated with pterostilbene (an homologous derivative of resveratrol) and further cultured under oxidative stress induced by uraemic serum (Chen et al., 2018). In addition, pretreatment of human HepG2 cells with the alkaloid epigoitrin increased SOD and GPx activity after exposure to H2O2 (Huo et al., 2017). All these responses found in cells have been associated to a general activation effect of antioxidant systems, in a complex scenario in which a battery of cytoprotective/defensive proteins including SOD, CAT and GPx are being up-regulated by the different compounds and treatments.
The analysis of transcriptional response of genes encoding the antioxidant enzymes and the further regulation of polypeptide translation has been less addressed. Unravelling this link is a key factor in order to understand the molecular mechanisms of cellular response against oxidative stress. For instance, a coordinated response in the induction of SOD, CAT and GPx activity, protein quantities and transcript amounts of CAT, SOD1 and GPx1 has been observed in HepG2 cells pretreated with the anthocyanin pelargonidin and then subjected to oxidative stress with citrinin (Babu et al., 2017). Similarly, human luteinized granulosa cells under oxidative stress induced by both H2O2 and dimethylfumarates exhibited a significant increase in SOD1 and CAT mRNAs and their respective polypeptides (Akino et al., 2018). In this work, a parallel and positive response in enzyme activities and transcripts for SOD (SOD1 and SOD2), CAT and GPx (GPx1) encoding genes in HSMMs cells as a consequence of TetraSOD® treatment was also found. Thus, the present results unravel the molecular basis of the cytoprotective effect of TetraSOD® in relation to the primary antioxidant enzymes.
In an attempt to go deeper in the knowledge of basal transcriptional regulation of antioxidant enzyme-encoding genes, the response to TetraSOD® treatment of genes NRF2 and HMOX-1 was then studied. NRF2 is a transcription factor widely expressed in eukaryotic cells which is known to activate antioxidant defenses. Actually, a number of reports have demonstrated that it is induced by antioxidant and chemopreventive compounds, which in turn regulates the expression of genes encoding both primary enzymes (that is, SOD, GPx, and CAT) and inducible phase II detoxifying enzymes such as HMOX1 (Krajka-KuzÌniak et al., 2015; Fetoni et al., 2015; Tebay et al., 2015; Babu et al., 2017; Bahar et al., 2017; Hong et al., 2017; Liu et al., 2017; Teixeira et al., 2017; Chen et al., 2018; Wang et al., 2018). The results have demonstrated up-regulation of both NRF2 and HMOX1 expression when HSMMs cells are incubated with TetraSOD®, which strongly suggests the participation of the NRF2/HMOX1 pathway in the increased expression of SOD, GPx and CAT genes, and hence in the protective antioxidant status induced by TetraSOD®. Additional regulation at translational level of primary antioxidant enzymes has not been addressed in this work, and hence it cannot be ruled out in the absence of more data.
Cellular protection against oxidative damage involves two types of antioxidants: “direct” antioxidants, which can undergo redox reactions and scavenge ROS intermediates, and “indirect” antioxidants, which are inducers of cytoprotective proteins (Dinkova-Kostova and Talalay, 2008). TetraSOD® exhibits a significantly high SOD activity (as well as GPx and CAT activities; not shown). The action of these enzymes might contribute to scavenging of extracellular ROS owing to their own antioxidant activities. But in addition, TetraSOD® treatment induces a key set of genes belonging to the antioxidant machinery present in eukaryotic cells. It has been suggested that the antioxidant actions observed after consumption of a SOD-enriched product elaborated from melon might be triggered by a cascade of events possibly involving the NRF2 pathway (Carillon et al., 2013a, b, 2014), but no further experimental supporting data could be provided. The present results with TetraSOD® fully agree with that hypothesis and represent an important contribution to the understanding of the more complex physiological protective and preventive effects derived from TetraSOD® consumption. However, further research will be necessary to unravel the whole mechanisms rendering such positive effects in vivo.
This study was sponsored by Fitoplancton Marino, S.L., the manufacturer of TetraSOD®. Authors disclose any financial/relevant interest that may have influenced this study.
REFERENCES
|
Akino N, Wada-Hiraike O, Terao H, Honjoh, H, Isono W, Fu H, Hirano M, Miyamoto Y, Tanikawa M, Harada M, Hirata T, Hirota Y, Koga K, Oda K, Kawana K, Fujii T, Osuga Y (2018). Activation of Nrf2 might reduce oxidative stress in human granulosa cells. Molecular and Cellular Endocrinology 470:96-104.
Crossref
|
|
|
|
Babu GRS, Anand T, Llaiyaraja N, Khanum F, Gopalan N (2017). Pelargonidin modulates Keap1/Nrf2 pathway gene expression and ameliorates citrinin-induced oxidative stress in HepG2 cells. Frontiers in Pharmacology 8:868.
Crossref
|
|
|
|
|
Bahar E, Kim JY, Yoon H (2017). Quercetin attenuates manganese-induced neuroinflammation by alleviating oxidative stress through regulation of apoptosis, iNOS/NF-κB and HO-1/Nrf2 Pathways. International Journal of Molecular Sciences 18:1989.
Crossref
|
|
|
|
|
Carillon J, Knabe L, Montalban A, Stévant M, Keophiphath M, Lacan D, Rouanet JM (2014). Curative diet supplementation with a melon superoxide dismutase reduces adipose tissue in obese hamsters by improving insulin sensitivity. Molecular Nutrition and Food Research 58:842-850.
Crossref
|
|
|
|
|
Carillon J, Romain C, Bardy G, Fouret G, Feillet-Coudray C, Gaillet S, Rouanet JM (2013b). Cafeteria diet induces obesity and insulin resistance associated with oxidative stress but not with inflammation: improvement by dietary supplementation with a melon superoxide dismutase. Free Radical Biology and Medicine 65:254-261.
Crossref
|
|
|
|
|
Carillon J, Rouanet JM, Cristol JP, Brion R (2013a). Superoxide dismutase administration, a potential therapy against oxidative stress related diseases: Several routes of supplementation and proposal of an original mechanism of action. Pharmaceutical Research 30(11):2718-2728.
Crossref
|
|
|
|
|
Chen ZW, Miu HF, Wang HP, Wu ZN, Wang WJ, Ling YJ, Xu XH, Sun HJ, Jiang X (2018). Pterostilbene protects against uremia serum-induced endothelial cell damage via activaction of Keap/Nrf2/HO-1 signaling. International Urology and Nephrology 50:559-570.
Crossref
|
|
|
|
|
Décordé K, Ventura E, Lacan D, Ramos J, Cristol JP, Rouanet JM (2010). An SOD rich melon extract Extramel® prevents aortic lipids and liver steatosis in diet-induced model of aterosclerosis. Nutrition, Metabolism and Cardiovascular Diseases 20:301-307.
Crossref
|
|
|
|
|
Dinkova-Kostova AT, Talalay P (2008). Direct and indirect antioxidant properties of inducers of cytoprotective proteins. Molecular Nutrition and Food Research 52:S128-S138.
Crossref
|
|
|
|
|
Espinosa-Diez C, Miguel V, Mennerich D, Kietzmann T, Sánchez-Pérez P, Cadenas S, Lamas S (2015). Antioxidant responses and celular adjustments to oxidative stress. Redox Biology 6:183-197.
Crossref
|
|
|
|
|
Fetoni AR, Paciello F, Rolesi R, Eramo SLM, Mancuso C, Troiani D, Paludetti G (2015). Rosmarinic acid up-regulates the noise-activated Nrf2/HO-1 pathway and protects against noise-induced injury in rat cochlea. Free Radical Biology and Medicine 85:269-281.
Crossref
|
|
|
|
|
Fiedor J, Burda K (2014). Potential role of carotenoids as antioxidants in human health and disease. Nutrients 6:466-488.
Crossref
|
|
|
|
|
Goyal MM, Basak A (2010). Human catalase: looking for complete identity. Protein Cell 1(10):888-897.
Crossref
|
|
|
|
|
Han YT, Han ZW, Yu GY, Wang YJ, Cui RY, Wang CB (2004). Inhibitory effect of polypeptide from Chlamys farreri on ultraviolet A-induced damage on human skin fibroblasts in vitro. Pharmacological Research 49:265-274.
Crossref
|
|
|
|
|
Hong C, Cao J, Wu CF, Kadioglu O, Schüffer A, Kauhl U, Klauck SM, Opatz T, Thines E, Paul NW, Efferth T (2017). The Chinese herbal formula Free and Easy Wanderer ameliorates oxidative stress through KEAP1-NRF2/HO-1 pathway. Scientific Reports 7:11551.
Crossref
|
|
|
|
|
Huo X, Liu C, Gao L, Xu X, Zhu N, Cao L (2017). Hepatoprotective effect of aqueous extract from the seeds of Orychophragmus violaceus against liver injury in mice and HepG2 cells. International Journal of Molecular Sciences 18:1197.
Crossref
|
|
|
|
|
Ighodaro OM, Akinloye OA (2018). First line defense antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defense grid. Alexandria Journal of Medicine 54:287-293.
Crossref
|
|
|
|
|
Jiao W, Wang Y, Kong L, Ou-yang T, Meng Q, Fu Q, Hu Z (2018). CART peptides activate the Nrf2/HO-1 antioxidant pathway and protect hippocampal neurons in a rat model of Alzheimer's disease. Biochemical and Biophysical Research Communications 501:1016-1022.
Crossref
|
|
|
|
|
Krajka-KuzÌniak V, Paluszczak J, Szaefer H, Baer-Dubowska W (2015). The activation of the Nrf2/ARE pathway in HepG2 hepatoma cells by phytochemicals and subsequent modulation of phase II and antioxidant enzyme expression. Journal of Physiology and Biochemistry 71:227-238.
Crossref
|
|
|
|
|
Letsiou S, Kalliampakou K, Gardikis K, Mantecon L, Infante C, Chatzikonstantinou M, Labrou NE, Flemetakis E (2017). Skin protective effects of Nannochloropsis gaditana extract on H2O2-stressed human dermal fibroblasts. Frontiers in Marine Science 4:221.
Crossref
|
|
|
|
|
Liao S, Qu Z, Li L, Zhou B, Gao M, Huang M, Li D (2018). HSF4 transcriptional regulates HMOX-1 expression in HLECs. Gene 655:30-34.
Crossref
|
|
|
|
|
Liu X, Liu H, Zhai Y, Li Y, Zhu X, Zhang W (2017). Laminarin protects against hydrogen peroxide-induced oxidative damage in MRC-5 cells possibly via regulating NRF2. Peer Journal 5:e3642.
Crossref
|
|
|
|
|
Livak KJ, Schmittgen TD (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2-(ΔΔCt) method. Methods 25:402-408.
Crossref
|
|
|
|
|
Margis R, Dunand C, Teixeira FK, Margis-Pinheiro M (2008). Glutathione peroxidase family - An evolutionary overview. FEBS Journal 275:3959-3970.
Crossref
|
|
|
|
|
Nelson SK, Bose SK, Grunwald GK, Myhill P, McCord JM (2006). The induction of human superoxide dismutase and catalase in vivo: A fundamentally new approach to antioxidant therapy. Free Radical Biology and Medicine 40:341-347.
Crossref
|
|
|
|
|
Putnam CD, Arvai AS, Bourne Y, Tainer JA (2000). Active and inhibited human catalase structures: ligand and NADPH binding and catalytic mechanism. Journal of Molecular Biology 296:295-309.
Crossref
|
|
|
|
|
Romao S (2015). Therapeutic value of oral supplementation with melon superoxide dismutase and wheat gliadin combination. Nutrition 31:430-436.
Crossref
|
|
|
|
|
Sheng Y, Abreu IA, Cabelli DE, Maroney MJ, Miller AF, Teixeira M, Valentine JS (2014). Superoxide dismutases and superoxide reductases. Chemical Reviews 114:3854-3918.
Crossref
|
|
|
|
|
Solano ME, Arck PC (2015). Heme-oxigenase 1: For better, for worse, in sickness and in health. Oncotarget 6(17):14733-14734.
Crossref
|
|
|
|
|
Stephenie S, Chang YP, Gnanasekaran A, Esa NM, Gnanaraj C (2020). An insight on superoxide dismutase (SOD) from plants for mammalian health enhancement. Journal of Functional Foods 68:103917.
Crossref
|
|
|
|
|
Tebay LE, Robertson H, Durant ST, Vitale SR, Penning TM, Dinkova-Kostova AT, Hayes JD (2015). Mechanisms of activation of the transcription factor Nrf2 by redox stressors, nutrient cues, and energy status and the pathways through which it attenuates degenerative disease. Free Radical Biology and Medicine 88:108-146.
Crossref
|
|
|
|
|
Teixeira TM, da Costa DC, Resende AC, Soulage CO, Bezerra FF, Daleprane JB (2017). Activation of Nrf2-antioxidant signaling by 1,25-dihydroxycholecalciferol prevents leptin-induced oxidative stress and inflammation in human endothelial cells. Journal of Nutrition 147(4):506-513.
Crossref
|
|
|
|
|
Tkachev VO, Menshchikova EB, Zenkov NK (2011). Mechanism of the Nrf2/Keap1/ARE signaling system. Biochemistry 76(4):407-422.
Crossref
|
|
|
|
|
Unamunzaga C, Mantecón E (2015). Patent "Method for obtaining a biomass of a microalga of the species Tetraselmis chuii enriched in superoxide dismutase (SOD)" European Patent Publication. Pub. No. EP 3292197 B1.
|
|
|
|
|
Vouldoukis I, Conti M, Krauss P, KamateÌ C, Blasquez S, Tefit M, Mazier D, Calenda A, Dugas B (2004). Supplementation with Gliadin-combined plant superoxide dismutase extract promotes antioxidant defences and protects against oxidative stress. Phytotherapy Research 18:957-962.
Crossref
|
|
|
|
|
Wang Y, Li L, Wang Y, Zhu X, Jiang M, Song E, Song Y (2018). New application of the commercial sweetener rebaudioside a as a hepatoprotective candidate: Induction of the Nrf2 signaling pathway. European Journal of Pharmacology 822:128-137.
Crossref
|
|
|
|
|
Zelko IN, Mariani TJ, Folz RJ (2002). Superoxide dismutase multigene family: A comparison of the CuZn-SOD (SOD1), MnSOD (SOD2), and EC-SOD (SOD3) gene structures, evolution, and expression. Free Radical Biology and Medicine 33(3):337-349.
Crossref
|
|
|
|
|
Zhou BR, Yin HB, Xu Y, Wu D, Zhang ZH, Yin ZQ, Permatasari F, Luo D (2012). Baicalin protects human skin fibroblasts from ultraviolet A radiation-induced oxidative damage and apoptosis. Free Radical Research 46(12):1458-1471.
Crossref
|
|