Full Length Research Paper

ABSTRACT
INTRODUCTION
In vitro androgenesis is a remarkable example of cell totipotency in plants (Reynolds, 1997; Góralski et al., 2005), and it can induce male gametophyte cells or their precursors to deviate their route toward a sporophytic program, resulting in the development of haploid embryos (Góralski et al., 1999; Bárány et al., 2005; Seguí-Simarro, 2010). Haploid plants are interesting in fundamental research owing to the development of sporophytes with the gametophytic number of chromosomes. Never-theless, their main practical utility is because of the generation of doubled haploid (DH) plants obtained by duplication of the chromosomes of haploid lineage either spontaneously or through induction by a microtubule-depolymerizing agent (Murovec and Bohanec, 2012). The DH generation is an exciting and powerful tool of pure line production (100% homozygous) for breeding programs (Kasha and Maluszynski, 2003; Dunwell, 2010; Germanà, 2010, 2011). Despite the knowledge obtained through model systems to study androgenesis such as in rapeseed (Brassica napus L.), tobacco (Nicotiana tabacum L.), or barley (Hordeum vulgare L.), the production of DH plants by androgenesis remains to be effectively standardized in tomato (Solanum lycopersicum, 2n = 2x = 24 chromosomes). The main hindrance in standardization is probably the recalcitrant nature of the crop (Bal and Abak, 2007; Seguí-Simarro and Nuez, 2007; Seguí-Simarro et al., 2011). S. lycopersicum has great economic importance in the horticultural industry (Wang et al., 2005), and it is considered as a good model for genetic studies of the Solanaceae family members (Hay et al., 2004; Koo et al., 2008).
The genotype dependency is a critical factor affecting the response in tomato androgenesis (Asoliman et al., 2007), and it has been reported that different cultivars of the same species exhibit diverse responses to another culture. For example, Zagorska et al. (1998) tested the androgenetic ability of 85 tomato cultivars, of which 53 were responsive and produced calli and a smaller number of cultivars developed regenerants. According to Seguí-Simarro and Nuez (2007), tomato genotypes can be divided into two categories: those inducible to form callus from meiocytes and those inducible to form embryos from microspores. Another critical aspect in androgenesis induction is the stage of microspore development at the time of anther excision and plating for in vitro culturing (Seguí-Simarro and Nuez, 2008a). In nearly all responsive crops, including model species such as rapeseed and tobacco, the inducible stage revolves around the first pollen mitosis, that is, from vacuolate microspore to early bicellular pollen (Touraev et al., 2001). In most tomato, the meiocyte is the most suitable developmental stage for androgenesis induction (Zamir et al., 1980; Summers et al., 1992; Shtereva et al., 1998; Brasileiro et al., 1999; Seguí-Simarro and Nuez, 2005; 2007). However, according to Seguí-Simarro and Nuez (2005), the stage with highest response in terms of callus generation in tomato is metaphase I to telophase II.
Hence, it is necessary to identify the appropriate anther developmental stage. Generally, traditional cytogenetic procedures such as squashing have been used for this purpose (Seguí-Simarro and Nuez, 2005). However, an accurate determination of microspore/pollen stage requires a complex cytogenetic analysis. Nevertheless, external morphological indicator, such as the corolla length, has been used because it is a simple procedure for measuring large-scale cases, although it is a less reliable method (Dunwell, 2010). An alternative method used to determine the microspore/pollen development stage is flow cytometry (FCM), which is a practical and efficient methodology for quantify (screening) nuclear DNA content. This technique allows analyzing the individual nuclei faster than the cytogenetic traditional methods. Moreover, this methodology can be also used for evaluating the DNA ploidy level of calli, and these results can be related to any effect on androgenetic potential of anthers. The loss of embryogenic and organogenic competence is one of the major problems encountered when developing in vitro regeneration systems for propagation and genetic manipulation of plants (Winkelmann et al.,1998).
Considering that different genotypes can trigger different androgenetic responses, the difficulty of selecting responsive anthers to the culture and solving why calli have not develop in embryogenic calli, the present study focused on (i) the introduction of a procedure to select microsporogenesis stages is potentially more responsive for callus induction, associating cytogenetic, and FCM technique; (ii) the use of a culture medium previously described as effective in the literature for callus induction in different genotypes; (iii) evaluation of responsiveness of different genotypes by callus production, and (iv) screening of the DNA ploidy level profile in the calli.
MATERIALS AND METHODS
Plant materials
Flower buds belonging to four genotypes of S. lycopersicum: ‘Debora’, ‘Santa Clara’, ‘Alambra’, and ‘Pizza Doro’ were provided by Dr. Derly J. H. da Silva, Vegetable Germplasm Bank Curator [Universidade Federal de Viçosa (UFV), Minas Gerais, Brazil]. Flower buds were collected in the morning at the experimental field of UFV; some of the collected buds were immediately fixed in a solution of methanol:acetic acid (3:1 v/v) for cytogenetic and FCM analyses, while the remaining were used for anther culturing. The fixative was changed three times, and the samples were stored at -20°C (Carvalho and Saraiva, 1997). The experiments were conducted at the Laboratório de Citogenética e Citometria, Departamento de Biologia Geral, UFV.
Monitoring of anther maturation by FCM
A total of 15 flower buds (1.0 to 6.0 mm) of all genotypes were removed from the donor plants in the morning, and their anthers were classified according to their size as small (0.5 to 0.7 mm), medium (0.8 to 1.5 mm), or large (1.6 to 1.9 mm). Four anthers of each bud were subjected to FCM procedure (total: 60 anthers) and one anther was reserved for cytogenetic analysis (total: 15 anthers). First, the anthers were softened by enzymatic maceration at 36°C as follows: the anthers were placed on a 70 µm mesh nylon filter submerged into a 2.0 mL microtube (Eppendorf) containing enzyme solution consisting of 2% cellulase (obtained from Trichoderma viride; Onozuka Yakult Honsha Co®), 1% cellulase (obtained from Aspergillus niger; Sigma®), 2.5% macerozyme (Yakult Honsha Co®), 2.5% pectinase powder (Sigma®), and 1% hemicellulase. The enzymatic maceration process was performed in two steps: the anthers were maintained in the enzymatic solution for 2 h and the softened anthers were macerated with a pin head so as to release the pollen mother cells (PMCs), followed by incubation of the PMCs in the same digestion solution for 30 min. Finally, the tubes were centrifuged for 5 min at 100 ×g, and the enzyme solution was discarded. Subsequently, the pellet was resuspended in 1 mL of OTTO-I lysis buffer (Otto, 1990) and the solution was homogenized with 12 pulses (approximately 1 s per pulse) by using a commercially available mini-handheld mixer to provide nuclei suspensions (Silva et al., 2010). The suspensions were filtered through 30 μm mesh nylon filter and centrifuged for 5 min at 100 ×g. The supernatant was discarded, and the pellet was resuspended in 100 μL of OTTO-I and incubated for another 10 min at the room temperature.
The nuclei suspension was stained with 1.5 mL of the OTTO-II buffer (Otto, 1990) supplemented with 1.5 mM 4′6′-diamino-2-phenylindole (DAPI; Sigma®), filtered through a 20 μm mesh nylon filter, and maintained in the dark for 30 min. The nuclei suspensions were analyzed by PAS-III flow cytometer (15-01-1000; Partec GmbH) equipped with a UV lamp (with emission at 378 nm), excitation filters (KG 1, BG 38, and UG 1), a GG 435 long-pass barrier filter for blue fluorescence, and a TK 420 nm dichroic mirror to supply epi-illumination. The equipment was carefully calibrated and aligned using microbeads and standard solutions as per the manufacturer’s recommendations. The FlowMax software (Partec GmbH) was used to process the data. The instrument gain was set up in such a way that the G0/G1 peak was positioned on channel 200 using nuclear suspension from leaves of the same plant.
Cytogenetic analysis
The anther that was separated in each bud was macerated in the same enzymatic solution used in the FCM analysis. Each anther was slightly fragmented with the tip of a scalpel blade in 50 µL of 45% acetic acid solution (v/v). The excessive material (wall anther) was removed from the slide and, on a hot plate, the solution containing the anther cell suspension was spread on the slide using a hair dryer. The slides were stained with 4.0 mM DAPI (Sigma®) and observed under the Olympus TM BX-60 fluorescence microscope.
Anther culture
Flower buds of S. lycopersicum genotypes ‘Debora’, ‘Santa Clara’, ‘Alambra’, and ‘Pizza Doro’ were sorted manually according to the size, from 1.0 to 6.0 mm at an interval of 1.0 mm. Next, the buds were pre-treated with 1% polyvinylpyrrolidone (PVP; Sigma®) solution in order to minimize the oxidation of the anthers and were maintained at 4°C for 48 h. After a cold shock, buds were sterilized under laminar flow hood with 70% (v/v) ethanol (Merck®) for 20 s, followed by immersion in 1.0% (v/v) solution of sodium hypochlorite (Merck®) for 5 min, and washed three times (3 min, each) with autoclaved distilled water (Zagorska et al., 1998). The anthers were aseptically removed; four anthers were inoculated into 60 × 15 mm Petri dishes (J. Prolab®) containing callus induction medium, and one anther was reserved for cytogenetic analysis to determine the stage of meiotic development. Induction medium consisted of MS salts and vitamins (Murashige and Skoog, 1962; Sigma®) supplemented with 2.8 g L−1 phytagel, 20 g L−1 sucrose, 0.1 g L−1 myo-inostol, 0.04 g L−1 cysteine, 1 mg L−1 6-(y,y-dimethylallylamino) purine (2iP; Sigma®) and 2 mg L−1 indole-3-acetic acid (IAA; Sigma®). The pH was adjusted to 5.7 prior to autoclaving. The growth regulators were filter-sterilized and added after autoclaving of the medium (adapted from Shtereva et al., 1998).
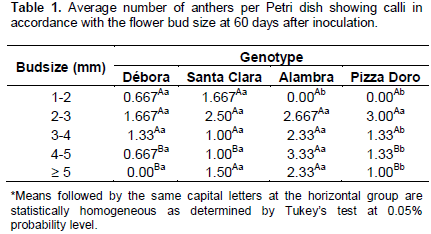
Petri dishes were sealed and kept in darkness for 1 month at 25 ± 2°C and subsequently transferred to a 16/8 h light/dark photoperiod regime with 36 µmol m−2 s−1 light radiation. Subcultures were made at 30 day intervals. The percentage of callus production was evaluated 60 days after inoculation. Anthers that did not respond to culture were discarded. The experiment followed a completely randomized design in a factorial 4 × 5, with a total of 20 treatments, corresponding to four genotypes and five bud sizes. Each treatment consisted of six Petri dishes containing four explants, and the parameter evaluated was the average of explants exhibiting calli. The data were submitted to analysis of variance (ANOVA) and Tukey’s test at 5% probability level, both performed by using the statistical program Genes (Cruz, 1997).
Calli screening by FCM
We analyzed the ploidy level of young calli (2 month old) by FCM, on six calli in each treatment, one per plate, except the plates where the anthers did not respond to induction of callus. Random pieces of each callus were removed and processed to release the nuclei, as described by Galbraith et al. (1983). Approximately 50 mg of callus was chopped in 100 µL of OTTO-I lysis buffer, followed by the addition of 400 µL of the same buffer. Then, the nuclei suspension was adjusted to 1 mL with the same buffer, filtered through 30 µm-mesh nylon filter into 2-mL microcentrifuge tubes, and centrifuged at 100 ×g for 5 min. The pellet was then incubated in 100 µL of OTTO-I lysis buffer for 10 min. The nuclei suspension was stained with 1.5 mL of the OTTO-II buffer supplemented with 1.5 mM DAPI (Sigma®), filtered through the 20 μm-mesh nylon filter, and maintained in the dark for 30 min. The suspension was analyzed by the Partec PAS® flow cytometer. For DNA ploidy level characterization of the calli, nuclear suspension from leaves of the correspondent plants (2C ploidy standard) was used to determine the G0/G1 peak. The peak relative to the 2C standard nuclei was set to channel 200. The histograms were obtained from the semi-logarithmic scale (log X-axis).
Cytological analysis
In order to confirm the nuclei isolation efficiency and to eliminate the possibility of nuclei aggregate formation, slides were prepared after the cytometric analysis. Nuclei suspensions were transferred to 2 mL of the microcentrifuge tubes and centrifuged at 100 ×g for 5 min. The supernatant was discarded, and 30 µL of the pellet were resuspended and then placed on a glass slide and covered with a coverslip. The images were captured by the DP71 digital camera coupled to the Olympus TM BX-60 fluorescence microscope using WU filter.
RESULTS
Monitoring of anther maturation by FCM and cytogenetic methods
For the first time, FCM approach was used to screen the meiotic progression in tomato anthers. Anthers of different sizes were evaluated by FCM in the four genotypes of tomato. The generated histograms showed coefficient of variation (CV) of < 5%, which is considered acceptable in plant FCM studies (Doležel and Bartoš, 2005). Histograms with low CV indicate that nuclei suspensions have intact, isolated, and stoichiometrically stained nuclei. Among the 60 anthers of different sizes analyzed, two histogram patterns were identified. The small anther profile consists of two histogram peaks, corresponding to the nuclei with 2C and 4C DNA content (Figure 1a). The large anther profile showed three peaks (1C, 2C and 4C ploidy level) (Figure 1b). Differences in the relative nuclear DNA content in PMCs and microspores within anthers were clearly evident from FCM analysis; young anthers (small) did not show n cells. In parallel to FCM analysis, cytogenetic analysis was performed to identify the meiotic phases correlated to the evaluated anther sizes. The developmental stages of microsporogenesis demonstrated that the (a) anthers of 0.5 to 0.7 mm size contained cells at interphase and prophase I; (b) anthers of 0.8 to 1.5 mm size contained meiocytes at metaphase I, anaphase I, telophase I, and meiosis II phases; and (c) anthers of > 1.6 mm size presented cells as tetrads and microspores (data not shown).
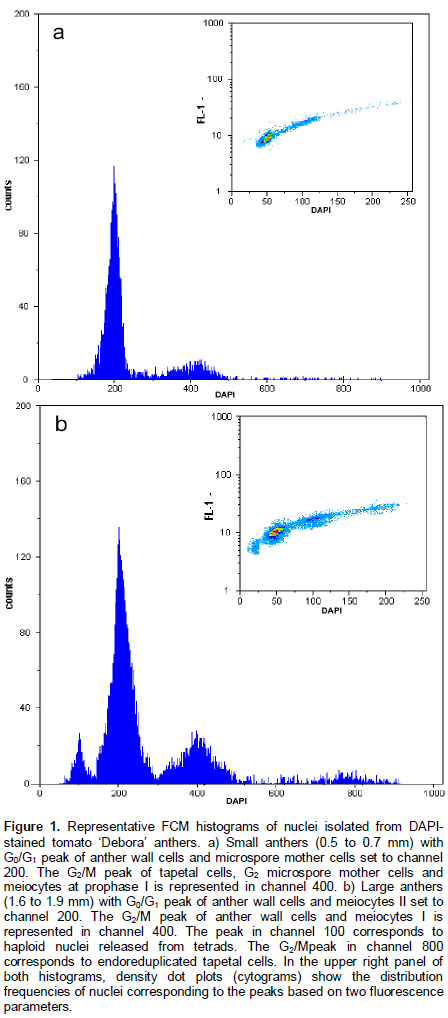
Anther culture
The four genotypes used in this study were tested for their androgenetic response by anther culture under the same experimental conditions. The FCM results in anther contributed to the choice of flower buds of the optimal size for inoculation of anthers, as suggested in the literature. Thus, anthers containing meiocytes (1.0 to 6.0 mm sized flower bud) were inoculated into the induction medium. Calli formation occurred 30 days after the beginning of each culturing. During the initial culturing, in the dark, the calli presented whitish color, which turned to green under 16/8 h photoperiod. Some shoots were formed on the generally compact calli (Figure 2). The evaluation of genotype × anther size interaction showed significant differences in some treatments for callus production, as can be visualized in Table 1. The callus production for ‘Pizza Doro’ genotype was influenced by the anther size (2 to 3 mm), and the genotype ‘Alambra’ influenced both 4 to 5 and > 5 mm anther lengths.

Calli screening by FCM
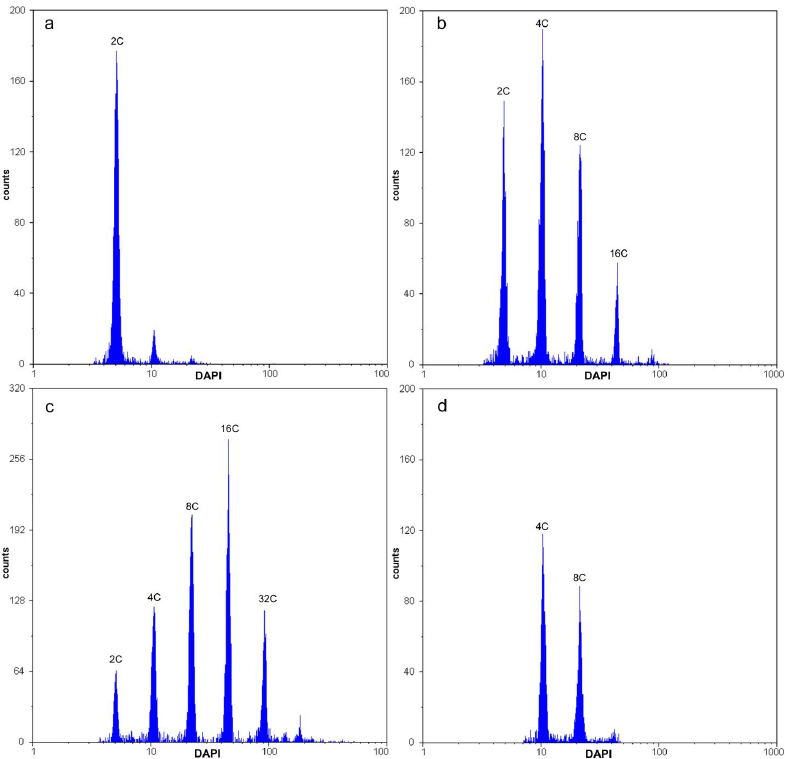
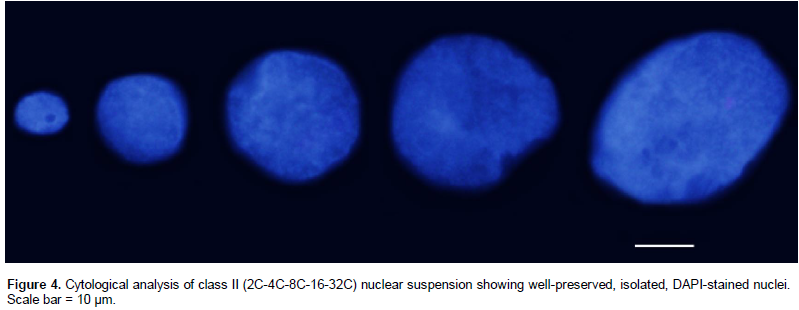
The procedure adopted here for nuclei extraction from calli was effective. The microscopic examination of nuclei suspensions revealed isolated nuclei, with only few nuclear aggregates. Moreover, the FCM analyses results were consistent in showing low CVs. Surprisingly, these FCM analyses showed that all calli were multiploid, a term suggested here to denote calli with a complex type of mixoploidy, presenting multiple DNA ploidy levels in the same calli and different ploidy levels in different calli. These multiploid calli were divided in five classes of DNA ploidy levels: class I (2C-4C-8C-16C), class II (2C-4C-8C-16C-32C), class III (4C-8C), class IV (4C-8C-16C-32C), and class V (8C-16C-32C) (Figure 3). We have presented ‘Débora’ FCM data to illustrate the multiploid event; however, the same classes were observed for the other genotypes as well. Each analyzed callus presented one of these classes. The calli showed the following distribution: 44.4% of the total belonging to class II, 27.78% to class I, 16.67% to class IV, and 5.55% to classes III and V. Class II was also analyzed cytologically to examine the nuclear size profile. Figure 4 illustrates five nuclear types in the ascending order of size that was compatible with five types of DNA ploidy levels as detected by FCM.
.png)
DISCUSSION
In this study, we show that flow cytometry can be a useful tool to screen the developmental stage of anthers. Brasileiro et al. (1999) also analyzed the correlation between tomato anther length and meiotic phase. These authors reported that flower buds of size 5.0 to 6.0 mm and anthers of size 2.0 to 2.3 mm showed cells in meiosis. Furthermore, Seguí-Simarro and Nuez (2005) observed that meiocytes were concentrated in the buds of length 4.0 to 5.9 mm. However, according to Zamir et al. (1980), anther and flower bud sizes are not directly correlated to the microsporogenesis stage. In the present study, different developmental stages were identified for similar bud sizes in the same plant, anthers in the same flower, and even within a single anther. Furthermore, the difference in the results of the present study and other literature reports can be attributed to the different genotypes analyzed in each experiment as well as the environmental and physiological conditions in which the plants were grown. The cytogenetic and FCM techniques allowed differentiation of the anthers at the interphase/ prophase and the tetrad/microspore stages. Small anthers at the interphase/prophase I displayed a histogram profile similar to that of the standard tomato leaf, whereas large anthers in the tetrad stage and microspore stages showed a peak corresponding to the haploid DNA content of microspores. Based on these results, flower buds of size 1.0 to 6.0 mm and anther size similar to that analyzed by FCM and cytogenetic procedures were inoculated into the induction medium.
The FCM analysis presented here represents a relatively simple, rapid, and reliable method to analyze and discriminate the development stage of anthers. The sample preparation in this study requires a small amount of anthers and the analysis is rapid. Moreover, a significant number of nuclei can be measured in a few minutes, which makes the results statistically robust. This procedure is interesting to accelerate the meiotic analysis, especially in plants in which no correlation exists between bud size or anthers with the meiotic stage. We have identified the response of four different tomato genotypes with different anther sizes in relation to calli production that were induced in an induction medium previously tested. Most studies on tomato anther culture have reported the occurrence of callogenesis ((Zamir et al., 1980; Shtereva et al., 1998; Brasileiro et al., 1999; Seguí-Simarro and Nuez, 2007; Corral-Martínez et al., 2005; 2011). However, considering the difficulty in achieving plant regeneration from calli in the test species besides the high recalcitrance of tomato to another culture (Seguí-Simarro and Nuez, 2005; Corral- Martínez et al., 2011), very little progress has been made in this context, with no standardized method being available for the generation of haploid and DH tomato plants (Seguí-Simarro and Nuez, 2007).
Various culture media have been tested for the induction of callus in tomato anthers. In this study, the induction medium suggested by Shtereva et al. (1998) was efficient for calli production in four genotypes of tomato anthers tested. Using the same medium, Asoliman et al. (2007) obtained a high frequency of somatic embryos from tomato anther culture, whereas Seguí-Simarro and Nuez (2005) obtained a high percentage of calli. Identification of the microsporogenic phase is considered as an important step for the successful anther culture. In this study, anthers were selected according to the size of flower buds and analyzed by FCM and cytogenetic procedures. In order to select the desired anther phase, Seguí-Simarro and Nuez (2005) and Góralski et al. (2005) identified the stage most responsive to androgenesis in tomato and maize plants, respectively, based on the relationship between in vitro anther development and cell characterization of the microspore/pollen. Our results show that anthers in meiosis, up to anaphase II, were equally responsive for callus induction.
Anthers containing microspores, obtained from buds with length ranging from 5.0 to 5.9 mm, did not respond to in vitro induction. Other studies corroborated that meiocytes is the most responsive stage for tomato anther culture (Zamir et al., 1980; Summers et al., 1992; Brasileiro et al., 1999; Seguí-Simarro and Nuez, 2005). Gresshoff and Doy (1972), Summers et al. (1992) and Brasileiro et al. (1999) reported that anthers showed higher frequencies of callus production during prophase I. Nevertheless, Seguí-Simarro and Nuez et al. (2005) identified callusogenic response in meiocytes from metaphase I until telophase II, whereas Shtereva et al. (1998) identified the response from prophase to telophase II as being optimal for tomato anther implantation. In most species, including model systems such as rapeseed and tobacco, the most sensitive period for inductive treatment is that near the first haploid mitosis, that is, from the vacuolated microspore to bicellular pollen stage (Touraev et al., 2001). Bal and Abak (2005) reported that the uninucleate stage was found to be suitable for the production of multicellular structure (proembryo) from tomato-isolated microspores. Occasional shoots were observed in this study, but these shoots did not regenerate into plants, probably owing to the recurrent polyploidization noted in calli. The ploidy was examined in an attempt to find out the reason of compact callus without embryogenic competence.
According to Ochatt et al. (2000), the increase in the DNA ploidy levels in calli can significantly complicate the regeneration of plants. The low efficiency of polyploidy calli regeneration is related to the loss of totipotency due to polyploidization (Colijn-Hooymans et al., 1994).
According to Phillips et al. (1994), changes in DNA ploidy levels are the most common somaclonal variations in tissue cultures. In fact, cell polyploidization in plant tissue culture has been described in numerous experimental systems. Park et al. (2010) cultivated Doritaenopsis leaf, root tip and somatic embryo in vitro in order to investigate their degrees of endoreduplication and found that somatic embryos showed the highest level (2C-64C). Several possible origins for somaclonal variations have been suggested, among which culture time is notable. Here, it was observed that this variable did not affect polyploidization in the calli, which presented populations of polyploid cells after two months of inoculation. In contrast, Salas et al. (2011) observed that culture time was the main factor influencing chromosome doubling in embryos derived from eggplant anther culture. Moreover, Song et al. (2000), Hao and Deng (2002) and Zhang et al. (2006) reported an increase in chromosome variations throughout subculturing.
The in vitro environment itself is an important source of stress. Though initial culture conditions promote cell proliferation, they may also cause defects in normal cytokinesis (Seguí-Simarro and Nuez, 2008b). In the present study, we used the culture medium suggested by Shtereva et al., (1998) assuming that the medium used successfully by the authors would be adequate for the genotypes used in this study. Thus, an interaction among growth regulators added to the culture medium and the genotypes of the donor plants may have induced endoreduplication cycles, resulting in high DNA ploidy levels in calli arising from tomato anther culture. It has been reported that the influence of growth regulators on calli induction depends on the donor plant genotype (Shtereva et al., 1998), representing one of the main factors that determine the type and frequency of genetic alterations (Jain, 2001). Seguí-Simarro and Nuez (2008a) also revealed that growth regulators induce increased DNA ploidy levels in microspores during anther culturing.
In our study, no population of haploid nuclei was observed in calli analysis by FCM, but multiploid nuclei were observed. According to Salas et al. (2011), this multiploidy could have somatic origin, as reported for anther culture of eggplant and the related species. In addition, Levenko et al. (1977) mentioned that cells of the anther wall contribute to the formation of mixoploid calli in tomato plants. Corral-Martínez et al. (2011) further showed that up to 83% of the calli produced from tomato anthers were derived from anther wall tissue. Nevertheless, Seguí-Simarro and Nuez (2007) and Corral-Martínez et al. (2011) also reported calli with haploid DNA content. Moreover, we suggested the possibility of the multiploidy to be meiotic cell lineage origin, whereas only anthers with meiocytes II were responsive. However, anthers with meiocytes after phase II possess somatic tissues capable of induction and which have not responded. This study allowed us to conclude that: (i) the association of cytogenetic and FCM techniques were effective in correlating the anther size with the meiotic developmental stage. Therefore, this method was considered to be adequate for the identification of the developmental stages of microsporo-genesis that are most responsive to callus induction on a large scale; (ii) Anthers containing meiocytes in the stages of prophase I up to anaphase II (2.0 to 3.9 mm) showed a higher percentage of callus formation. However, (iii) no haploid material was obtained in all genotypes tested; (iv) multiploid calli were identified in the short-term culture (two months), suggesting that this variable (time of culturing) was not directly responsible for recurrence of endoreduplication. The occurrence of these endoreduplication cycles in tomato anther culture has been reported for the first time in the present paper. Most reports of anther culture reported that the DNA ploidy levels were not exceptionally high. Since we used different genotypes in a culture medium considered suitable for haploid production, our results indicate a greater importance of the interaction between genotype and growth regulators in the process of obtaining haploids that promoted calli with different and high DNA ploidy levels.
CONFLICT OF INTERESTS
The authors did not declare any conflict of interest.
ACKNOWLEDGEMENTS
We would like to thank Conselho Nacional de Pesquisa (CNPq, Brazil), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES, Brazil), and Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG, Brazil) for the financial support to our study.
REFERENCES
| Asoliman SS, Ismail TA, Zaki MA, Amer ES (2007). Genetic studies on tomato anther. Factors affecting induction of androgenesis in tomato anther culture. Afr. Crop Sci. Conf. Proc. 8: 759-768 | ||||
|
Bal U, Abak K (2005). Induction of symmetrical nucleus division and multicellular structures from the isolated microspores of Lycopersicon esculentum. Mill Biotechnol. Biotechnol. Equipment 19:35-42. Crossref |
||||
|
Bal U, Abak K (2007). Haploidy in tomato (Lycopersicon esculentum Mill.): a critical review. Euphytica 158:1-9. Crossref |
||||
|
Bárány I, González-Melendi P, Fadon B, Mitykó J, Risue-o MC, Testillano PS (2005). Microspore-derived embryogenesis in pepper (Capsicum annuum L.): subcellular rearrangements through development. Biol. Cell 97:709-722. Crossref |
||||
|
Brasileiro ACR, Willadino L, Guerra M, Colaço W, Meunier I, Camara TR (1999). Efeitos do estádio de desenvolvimento da antera e da radiação gama na formação de calos derivados de anteras de tomate. Sci. Agric. 56: 835-842. Crossref |
||||
|
Carvalho CR, Saraiva L (1997). High-Resolution HKG-Banding in Maize Mitotic Chromosomes. J. Plant Res. 110:417-420 Crossref |
||||
|
Colijn-Hooymans CM, Hakkert JC, Jansen J, Custers JBM (1994). Competence for regeneration of cucumber cotyledons is restrict to specific developmental stages. Plant Cell Tissue Organ Cult. 39:211-217. Crossref |
||||
|
Corral-Martínez P, Nuez F, Seguí-Simarro JM (2011). Genetic, quantitative and microscopic evidence for fusion of haploid nuclei and growth of somatic calli in cultured ms1035 tomato anthers. Euphytica 178: 215-228. Crossref |
||||
| Cruz CD (1997). Programa GENES—Aplicativo Computacional em Genética e Estatística. Editora UFV, Viçosa, MG, p. 442 | ||||
|
Doležel J, Bartoš J (2005). Plant DNA flow cytometry and estimation of nuclear genome size. Ann. Bot. 95: 99-110. Crossref |
||||
|
Dunwell JM (2010). Haploids in flowering plants: origins and exploitation. Plant Biotechnol. J. 8:377-424. Crossref |
||||
|
Galbraith DW, Harkins KR, Maddox JM, Ayres NM, Sharma DP, Firoozabady E (1983). Rapid flow cytometric analysis of the cell cycle in intact plant tissues. Science 220: 1049-1051 Crossref |
||||
|
Germanà MA (2010). Anther culture for haploid and doubled haploid production. Plant Cell Tissue Organ Cult. 104: 283-300. Crossref |
||||
|
Germanà MA (2011). Gametic embryogesis and haploid technology as valuable support to plant breeding. Plant Cell Rep. 30: 839-857. Crossref |
||||
| Góralski G, Matthys-Rochon E, Vergne P, Przywara L (1999). Androgenic development : a fascinating embryo formation process. Acta Bio. Crac. 41:51-65 | ||||
| Góralski G, Rozier F, Matthys-Rochon E, Przywara L (2005). Cytological features of various microspore derivatives appearing during culture of isolated maize microspores. Acta Biol. Crac. Ser. Bot. 47:75-83 | ||||
|
Gresshoff P, Doy C (1972). Development and differentiation of haploid Lycopersicon esculentum (tomato). Planta 107:161-170. Crossref |
||||
|
Hao Y-J, Deng X-X (2002). Occurrence of chromosomal variations and plant regeneration from long-term-cultured citrus callus. In Vitro Cell. Dev. Biol. - Plant 38:472-476. Crossref |
||||
|
Hay A, Craft J, Tsiantis M (2004). Plant hormones and homeoboxes: bridging the gap? BioEssays 26: 395-404. Crossref |
||||
|
Jain SM (2001). Tissue culture-derived variation in crop improvement. Euphytica 118: 153-166. Crossref |
||||
|
Kasha KJ, Maluszynski M (2003). Production of doubled haploids in crop plant. An introduction. In: Maluszynski M, Kasha KJ, Forster BP, Szarejko I (eds) Doubled haploid production in crop plants. A manual. Kluwer Academic Publishers, Dordretch, the Netherlands, pp. 1-4. Crossref |
||||
|
Koo D-H, Jo S-H, Bang J-W, Park H-M, Lee S, Choi D (2008). Integration of cytogenetic and genetic linkage maps unveils the physical architecture of tomato chromosome 2. Genetics 179: 1211-1220. Crossref |
||||
| Levenko BA, Kunakh VA, Yurkova GN (1977). Studies on callus tissue from anthers. I. Tomato. Phytomorphology 27: 377-383 | ||||
|
Murashige T, Skoog F (1962). A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 15: 473-479 Crossref |
||||
|
Murovec J, Bohanec B (2012). Haploids and doubled haploids in plant breeding. In: Abdurakhmonov I (ed) Plant Breeding. In Tech Europe, Rijeka, pp. 87-106. Crossref |
||||
|
Ochatt SJ, Mousset-Déclas C, Rancillac M (2000). Fertile pea plants regenerate from protoplasts when calluses have not undergone endoreduplication. Plant Sci. 156:177-18. Crossref |
||||
|
Otto FJ (1990). DAPI staining of fixed cells for high-resolution flow cytometry of nuclear DNA. In: Darzynkiewiez Z, Crissman HA, Robinson JP (eds) Methods in Cell Biology. Academic Press, San Diego, pp. 105-110. Crossref |
||||
|
Park S-Y, Edward CY, Paek K-Y (2010). Endoreduplication in Phalaenopsis is affected by light quality from light-emitting diodes during somatic embryogenesis. Plant Biotechnol. Rep. 4:303-309. Crossref |
||||
|
Phillips RL, Kaepler SM, Olhoft P (1994). Genetic instability of plant tissue culture: breakdown if normal controls. Proc. Natl. Acad. Sci. USA 91:5222-5226. Crossref |
||||
|
Reynolds TL (1997). Pollen embryogenesis. Plant Mol. Biol. 33: 1-10 Crossref |
||||
|
Salas P, Prohens J, Seguí-Simarro JM (2011). Evaluation of androgenic competence through anther culture in common eggplant and related species. Euphytica 182: 261-274. Crossref |
||||
|
Seguí-Simarro JM (2010). Androgenesis revisited. Bot. Rev. 76:377-404. Crossref |
||||
|
Seguí-Simarro JM, Corral-Martínez P, Parra-Vega V, González-García B (2011). Androgenesis in recalcitrant solanaceous crops. Plant Cell Rep. 30:765-778. Crossref |
||||
|
Seguí-Simarro JM, Nuez F (2005). Meiotic metaphase I to telophase II as the most responsive stage during microspore development for callus induction in tomato (Solanum lycopersicum) anther cultures. Acta Physiol. Plant 27: 675-685. Crossreff |
||||
|
Seguí-Simarro JM, Nuez F (2007). Embryogenesis induction, callogenesis, and plant regeneration by in vitro culture of tomato isolated microspores and whole anthers. J. Exp. Bot. 58:1119-1132. Crossref |
||||
|
Seguí-Simarro JM, Nuez F (2008a). How microspores transform into haploid embryos: changes associated with embryogenesis induction and microsporederived embryogenesis. Physiol. Plant. 134:1-12. Crossref |
||||
|
Seguí-Simarro JM, Nuez F (2008b). Pathways to doubled haploidy: chromosome doubling during androgenesis. Cytogenet. Genome Res. 120:358-369. Crossref |
||||
|
Shtereva LA, Zagorska NA, Dimitrov BD, Kruleva MM, Oanh HK (1998). Induced androgenesis in tomato (Lycopersicon esculentum Mill). II. Factors affecting induction of androgenesis. Plant Cell Rep. 18:312-317. Crossref |
||||
|
Silva TCR, Abreu IS, Carvalho CR (2010). Improved and reproducible flow cytometry methodology for nuclei isolation from single root meristem. J. Bot. 2010:1-7. Crossref |
||||
| Song X, Xia G, Chen H (2000). Chromosomal variation in long-term cultures of several related plants in Triticinae. Acta Phytobiol. Sin. 26: 33-38 | ||||
| Summers WL, Jaramillo J, Bailey T (1992). Microspore developmental stage and anther length lnfluence the induction of tomato anther callus. HortScience 27: 838-840 | ||||
|
Touraev A, Pfosser M, Heberle-Bors E (2001). The microspore: a haploid multipurpose cell. Adv. Bot. Res. 35: 53-109 Crossref |
||||
| Wang XF, Knoblauch R, Leist N (2005). Varietal discrimination of tomato (Lycopersicon esculentum L.) by ultrathin-layer isoelectric focusing of seed protein. Seed Sci. Technol. 28: 521-526 | ||||
|
Winkelmann T, Sangwan RS, Schwenkel HG (1988). Flow cytometric analyses in embryogenic and non-embryogenic callus lines of Cyclamen persicum Mill.: relation between ploidy level and competence for somatic embryogenesis. Plant Cell Rep. 17:400-404. Crossref |
||||
|
Zagorska NA, Shtereva LA, Dimitrov BD, Kruleva MM (1998). Induced androgenesis in tomato (Lycopersicon esculentum Mill.) - I. Influence of genotype on androgenetic ability. Plant Cell Rep. 17:968-973. Crossref |
||||
|
Zamir D, Jones RA, Kedar N (1980). Anther culture of male-sterile tomato (Lycopersicon esculentum Mill.) mutants. Plant Sci. Lett. 17:353-361. Crossref |
||||
|
Zhang J-E, Guo W-W, Deng X-X (2006). Relationship between ploidy variation of citrus calli and competence for somatic embryogenesis. Acta Genet. Sin. 33: 647-654. Crossref |
||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


