Full Length Research Paper

ABSTRACT
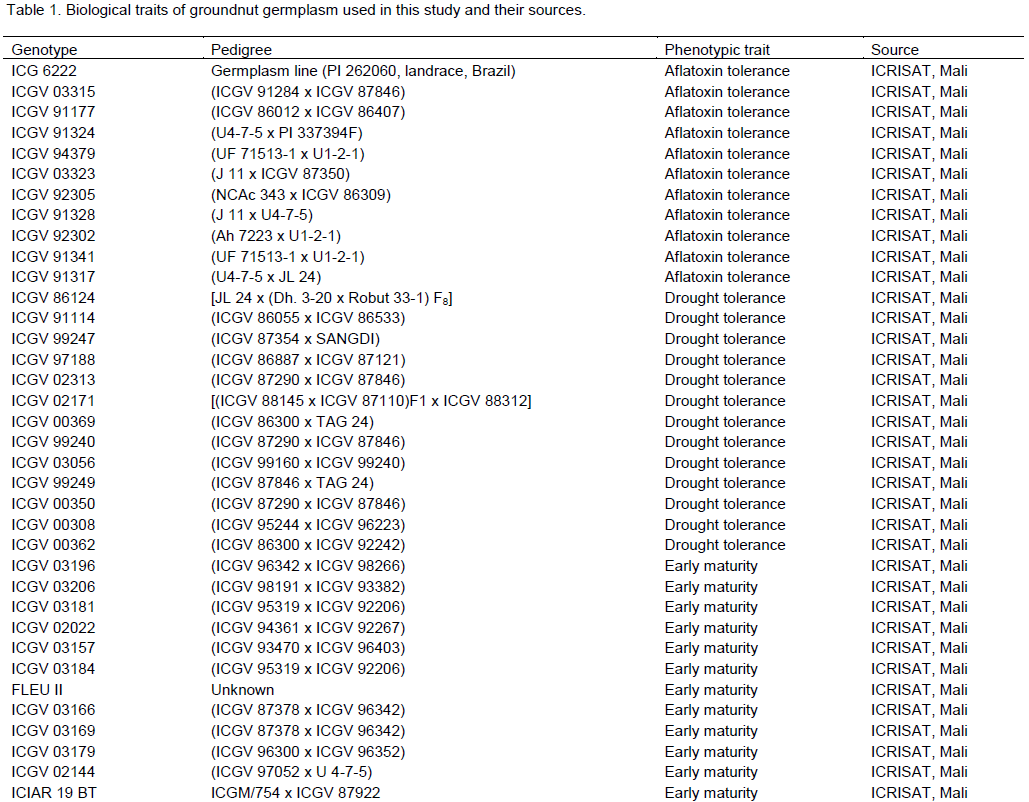
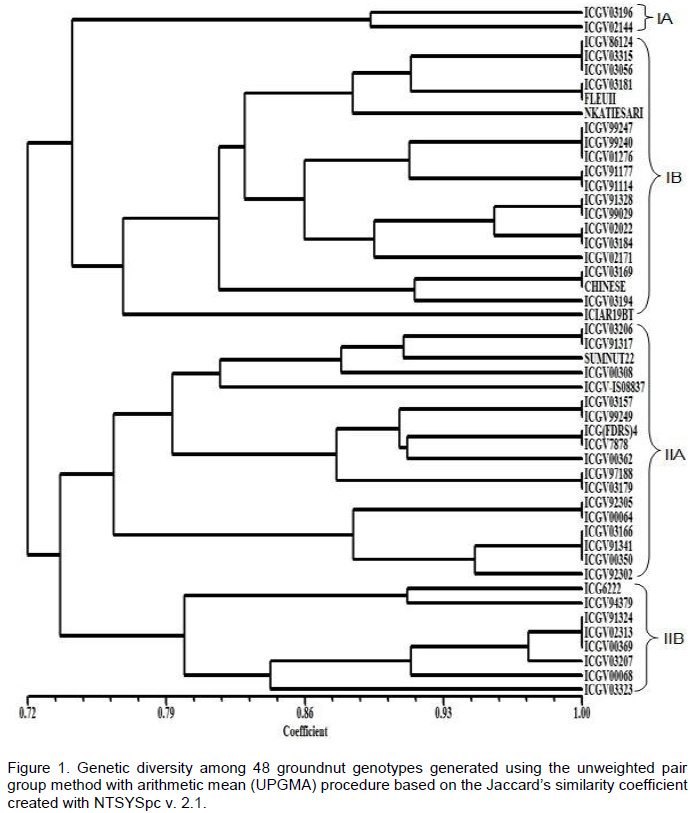
Groundnut (Arachis hypogaea L.) is the most important grain legume in Ghana. However, its production is constrained by a myriad of biotic and abiotic stresses which necessitate the development and use of superior varieties for increased yield. Germplasm characterisation both at the phenotypic and molecular level is important in all plant breeding programs. The aim of this study was to characterise selected advanced breeding groundnut lines with different phenotypic attributes at the molecular level using simple sequence repeats (SSR) markers in Ghana. A total of 53 SSR markers were screened and 25 were found to be polymorphic with an average polymorphic information content (PIC) value of 0.57. Of the 48 groundnut genotypes studied, 67% showed very close relationship (~100% similarity) with one or more genotypes among themselves. In fact, there were 14 instances where two to three genotypes within the same sub-cluster exhibited 100% similarity even though they displayed different phenotypic attributes. The remaining 33% of the groundnut genotypes were distant from each other and could therefore serve as effective parental material for future work. In this study, the SSR-based markers were found to be quite discriminatory in discerning variations between and among groundnut lines even where the level of variation was low. Microsatellite-based markers therefore represent a useful tool for dissecting genetic variations in cultivated crops, especially groundnut.
Key words: Phenotypic traits, DNA extraction, PCR amplification, simple sequence repeats (SSR) markers, alleles, polymorphic information content, Jaccard’s similarity coefficient.
Abbreviation: PIC, Polymorphic information content; RFLP, restriction fragment length polymorphism; AFLP, amplified fragment length polymorphism; SCARs, sequence characterised amplified regions; RAPDs, random amplified polymorphic DNA; SSR, simple sequence repeats SNP, single nucleotide polymorphism.INTRODUCTION
MATERIALS AND METHODS

RESULTS AND DISCUSSION

.png)
CONFLICT OF INTERESTS
The authors did not declare any conflict of interest.
ACKNOWLEDGEMENTS
This study was supported with funds from the Bill and Melinda Gates Foundation (BMGF) under the auspices of the BMGF-Project on Capacity Building in Africa (awarded to Tshwane University of Technology, Pretoria). The authors are grateful to the Foundation for a doctoral fellowship awarded under the BMGF-Project, to CSIR-Savanna Agricultural Research Institute, Ghana, for grant of a study leave, and to ICRISAT for seed material and training support. The DST/NRF South African Research Chair in Agrochemurgy and Plant Symbioses and the Tshwane University of Technology are duely acknowledged for their continued funding support of FDD’s research.
REFERENCES
|
Abubakari AH, Nyarko G, Yidana JA, Mahunu GK, Abagale FK, Quainoo A, Chimsah F, Avornyo V (2012). Comparative studies of soil characteristics in Shea parklands of Ghana. J. Soil Sci. Environ. Manag. 3: 84-90. |
|
|
Asibuo JY, Akromah R, Safo-Kantanka O, Adu-Dapaah HK, Ohemeng-Dapaah S, Agyeman A(2008). Chemical composition of groundnut, Arachis hypogaea (L) landrace. Afr. J. Biotechnol. 7: 2203-2208. |
|
|
Barkley NA, Wang ML, Pittman RN (2011). A real-time PCR genotyping assay to detect FAD2A SNPs in peanuts (Arachis hypogaea L.). Electron. J. Biotechnol. 14: 1-9. |
|
|
Carvalho MA, Quesenberry KH, Gallo M(2010). Comparative assessment of variation in the USA Arachispintoi (Krap. and Greg.) germplasm collection using RAPD profiling and tissue culture regeneration ability. Plant Syst. Evol. 288: 245-251. |
|
|
Cuc LM, Mace ES, Crouch JH, Quang VD, Long TD, Varshney RK (2008). Isolation and characterization of novel microsatellite markers and their application for diversity assessment in cultivated groundnut (Arachishypogaea L.). BMC Plant Biol. 8:55. |
|
|
Dakora FD, Aboyinga RA, Mahama Y, Apaseku J (1987). Assessment of N2 fixation in groundnut (Arachis hypogaea L.) and cowpea (Vigna unguiculata L. Walp.) and their relative N contribution to a succeeding maize crop in Northern Ghana. Mircen J. Appl. Microbiol. Biotechnol. 3: 389-399. |
|
|
FAO (2014). FAOSTAT. |
|
|
Garcia GM, Stalker HT, Shroeder E, Kochert G (1996). Identification of RAPD, SCAR and RFLP markers tightly linked to nematode resistance genes introgressed from Arachis cardenasii to Arachis hypogaea. Genome 39: 836-845. |
|
|
Gautami B, Pandey MK, Vadez V, Nigam SN, Ratnakumar P, Krishnamurthy L, Radhakrishnan T, Gowda MVC, Narasu ML, Hoisington DA, Knapp SJ, Varshney RK (2012). Quantitative trait locus analysis and construction of consensus genetic map for drought tolerance traits based on three recombinant inbred line populations in cultivated groundnut (Arachis hypogaea L.). Mol. Breed. 30: 757-772. |
|
|
Guo Y, Khanal S, Tang S, Bowers JE, Heesacker AF, Khalilian N, Nagy ED, Zhang D, Taylor CA, Stalker HT, Ozias-Akins P, Knapp SJ (2012). Comparative mapping in intraspecific populations uncovers a high degree of macrosynteny between A- and B-genome diploid species of peanut. BMC Genomics 13:608. |
|
|
He G, Prakash CS(1997). Identification of polymorphic DNA markers in cultivated peanut (Arachis hypogaea L.). Euphytica 97: 143-149. |
|
|
Holbrook CC, Stalker HT (2003). Peanut Breeding and Genetic Resources, In: Janick, J. (Ed.), Plant Breeding Reviews. John Wiley & Sons, Inc., pp. 297-356. |
|
|
Hopkins MS, Casa AM, Wang T, Mitchell SE, Dean RE, Kochert G, Kresovich S (1999). Discovery and characterization of polymorphic simple sequence repeats (SSRs) in cultivated peanut (Arachis hypogaea L.). Crop Sci. 39: 1243-124. |
|
|
Janila P, Nigam SN, Pandey MK, Nagesh P, Varshney RK (2013). Groundnut improvement: use of genetic and genomic tools. Front. Plant Sci. 4: 1-16. |
|
|
Jiang H, Liao B, Ren X, Lei Y, Mace ES, Fu T, Crouch JH (2007). Comparative assessment of genetic diversity of peanut (Arachis hypogaea L.) genotypes with various levels of resistance to bacterial wilt through SSR and AFLP analyses. J. Genet. Genomics 34: 544-554. |
|
|
Kochert G, Halward T, Branch WD, Simpson CE (1991). RFLP variability in peanut (Arachis hypogaea L.) cultivars and wild species. Theor. Appl. Genet. 81: 565-570. |
|
|
Liu K, Muse SV (2005). PowerMarker: Integrated analysis environment for genetic marker data. Bioinformatics 21: 2128-2129. |
|
|
Mace ES, Buhariwalla HK, Crouch JH (2003). A high-throughput DNA extraction protocol for tropical molecular breeding programs. Plant Mol. Biol. Rep. 21: 459a-459h. |
|
|
Mace ES, Phong DT, Upadhyaya HD, Chandra S, Crouch JH (2006). SSR analysis of cultivated groundnut (Arachishypogaea L.) germplasm resistant to rust and late leaf spot diseases. Euphytica 152:317-330. |
|
|
Martey E, Wiredu AN, Oteng-Frimpong R (2015). Baseline study of groundnut in Northern Ghana. LAP Lambert Academic Publishing. |
|
|
MoFA-SRID (2011). Agriculture in Ghana: Facts and Figures. Minist. Food Agric. Ghana, Stat. Res. Inf. Dir. 1-53. |
|
|
Mokgehle SN, Dakora FD, Mathews C (2014). Variation in N2 fixation and N contribution by 25 groundnut (Arachis hypogaea L.) varieties grown in different agro-ecologies, measured using 15N natural abundance. Agric. Ecosyst. Environ. 195: 161-172. |
|
|
Mondal S, Badigannavar AM (2010). Molecular diversity and association of SSR markers to rust and late leaf spot resistance in cultivated groundnut (Arachis hypogaea L.). Plant Breed. 129: 68-71. |
|
|
Naab JB, Seini SS, Gyasi KO, Mahama GY, Prasad PVV, Boote KJ, Jones JW (2009). Groundnut yield response and economic benefits of fungicide and phosphorus application in farmer-managed trials in northern Ghana. Exp. Agric. 45: 385-399. |
|
|
Naab JB, Tsigbey FK, Prasad P, Boote KJ, Bailey J, Brandenburg RL (2005). Effects of sowing date and fungicide application on yield of early and late maturing peanut cultivars grown under rainfed conditions in Ghana. Crop Prot. 24: 325-332. |
|
|
Nagy ED, Guo Y, Tang S, Bowers JE, Okashah RA, Taylor CA, Zhang D, Khanal S, Heesacker AF, KhalilianN, Farmer AD, Carrasquilla-Garcia N, Penmetsa RV, Cook DR, Stalker HT, Nielsen N, Ozias-Akins P, Knapp SJ (2012). A high-density genetic map of Arachis duranensis, a diploid ancestor of cultivated peanut. BMC Genomics 13:469. |
|
|
Nigam SN (2000). Some strategic issues in breeding for high and stable yield in groundnut in India. J. Oilseeds Res. 17: 1-10. |
|
|
Nyemba RC, Dakora FD (2010). Evaluating N2 fixation by food grain legumes in farmers' fields in three agro-ecological zones of Zambia, using 15N natural abundance. Biol. Fertil. Soils 46: 461-470. |
|
|
Padi FK (2008). Genotype × Environment interaction for yield and reaction to leaf spot infections in groundnut in semiarid West Africa. Euphytica 164: 143-161. |
|
|
Pandey MK, Gautami B, Jayakumar T, Sriswathi M, Upadhyaya HD, Gowda MVC, Radhakrishnan T, Bertioli DJ, Knapp SJ, Cook DR, Varshney RK (2012a). Highly informative genic and genomic SSR markers to facilitate molecular breeding in cultivated groundnut (Arachis hypogaea L.). Plant Breed. 131: 139-147. |
|
|
Pandey MK, Monyo E, Ozias-Akins P, Liang X, Guimarães P, Nigam SN, Upadhyaya HD, Janila P, Zhang X, Guo B, Cook DR, Bertioli DJ, Michelmore R, Varshney RK (2012b). Advances in Arachis genomics for peanut improvement. Biotechnol. Adv. 30: 639-651. |
|
|
Perrier X, Jacquemound-Collet JP (2006). DARwin Software. |
|
|
Rademacher-Schulz C, Schraven B, Mahama ES (2014). Time matters: shifting seasonal migration in Northern Ghana in response to rainfall variability and food insecurity. Clim. Dev. 6: 46-52. |
|
|
Rakoczy-Trojanowska M, Bolibok H (2004). Characteristics and a comparison of three classes of microsatellite-based markers and their application in plants. Cell. Mol. Biol. Lett. 9: 221-238. |
|
|
Ravi K, Vadez V, Isobe S, Mir RR, Guo Y, Nigam SN, Gowda MVC, Radhakrishnan T, Bertioli DJ, Knapp SJ, Varshney RK (2011). Identification of several small main-effect QTLs and a large number of epistatic QTLs for drought tolerance related traits in groundnut (Arachis hypogaea L .). Theor. Appl. Genet. 122: 1119-1132. |
|
|
Rohlf FJ (1992). NTSYS-PC: numerical taxonomy and multivariate analysis system. Exeter Software, New York. |
|
|
Shirasawa K, Bertioli DJ, Varshney RK, Moretzsohn MC, Leal-Bertioli SCM, Thudi M, Pandey MK, Rami J-F, Foncéka D, Gowda MVC, Qin H, Guo B, Hong Y, Liang X, Hirakawa H, Tabata S, Isobe S (2013). Integrated consensus map of cultivated peanut and wild relatives reveals structures of the A and B genomes of Arachis and divergence of the legume genomes. DNA Res. 20: 173-184. |
|
|
Shoba D, Manivannan N, Vindhiyavarman P (2010). Genetic diversity analysis of groundnut genotypes using SSR markers. Electron. J. Plant Breed. 1: 1420-1425. |
|
|
Sujay V, Gowda MVC, Pandey MK, Bhat RS, Khedikar YP, Nadaf HL, Gautami B, Sarvamangala C, Lingaraju S, Radhakrishnan T, KnappSJ, Varshney RK (2012). Quantitative trait locus analysis and construction of consensus genetic map for foliar disease resistance based on two recombinant inbred line populations in cultivated groundnut (Arachis hypogaea L.). Mol. Breed. 30: 773-788. |
|
|
Tang R, Gao G, He L, Han Z, Shan S, Zhong R, Zhou C, Jiang J, Li Y, Zhuang W (2007). Genetic diversity in cultivated groundnut based on SSR markers. J. Genet. Genomics 34: 449-459. |
|
|
Tsigbey FK, Brandenburg RL, Clottey VA (2003). Peanut production methods in northern Ghana and some disease perspectives. Online J. Agron. 34: 36-47. |
|
|
Wang CT, Yang XD, Chen DX, Yu SL, Liu GZ, Tang YY, Xu JZ (2007). Isolation of simple sequence repeats from groundnut. Electron. J. Biotechnol. 10: 473-480. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


