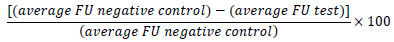ABSTRACT
Agriculture suffers considerable losses in its production caused by plant diseases. In the citrus culture some problems were associated with citrus canker caused by Xanthomonas citri subsp. citri. That same way, sugarcane plantations are impaired to leaf scald disease caused by Xanthomonas albilineans. One of the measures used to contain citrus canker is the spray of cupric bactericides; however for scalding leaves there are no satisfactory control. Due to the phytosanitary problem caused by Xanthomonas (X. citri and X. albilineans) and by the difficulty of its control, the search for new forms of defense, less harmful to the environment, has become increasingly required. Thus, the aim of this work was to obtain crude organic extracts from filamentous fungi isolated from Antarctic soil samples and assess its bioactivity potential against X. citri and X. albilineans. One hundred and twenty-two extracts were tested, seven extracts inhibited the cell growth of X. citri, one was bioactive only against X. albilineans and one extract inhibited the cell growth of both bacteria. The bioactive extracts had a mean inhibition value of 96% against both bacteria. The values of MIC90 and MBC of bioactive extracts were also determined; for X. citri the isolate 3.1 Fe presented the lowest values of MIC90 (0.28 mg/mL) and MBC (1.0 mg/mL) and the two bioactive isolates for X. albilineans (1.1-Fe and B-Fe) presented the same values of MIC90 (1.4 mg/mL) and MBC (1.5 mg/mL). The filamentous fungi that produced positive extracts were identified as belonging to the genus Pseudogymnoascus (n=8) and Cladosporium (n=1). The filamentous fungi isolated from Antarctic soil produced compounds with bioactivity against phytopathogens from the Xanthomonas genus.
Key words: Antarctic fungi; antibacterial action; citrus canker; leaf scald; natural extracts.
Xanthomonas are rod-shaped Gram-negative and obligate aerobe bacteria, which have a polar flagellum (Tessmann, 2002). This genus comprises 27 species that cause serious disease in about 400 plants hosts (Ryan et al., 2011) and affect at least 124 monocotyledonous and 268 dicotyledonous plant species (Leyns et al., 1984).
All citrus species and varieties of commercial importance are affected by citrus canker caused by Xanthomonas citri subsp. citri (Schaad et al., 2006). Every year orchards around the world suffer losses due to citrus canker, mainly countries with favorable climates for the development of X. citri. The control of this disease is accomplished by uses of less susceptible varieties, eradication of contaminated plants, decontamination of seeds and equipment, use of windbreakers and mostly use of antibiotics and copper sprays. This causes damage to the environment due to its accumulative potential in soil and trophic chain (Rocha and Azevedo, 2017).
The leaf scald in sugarcane is a disease caused by X. albilineans. This infection can be characterized in three ways: Asymptomatic or latent form (plants can tolerate the pathogen for a few weeks without exhibiting symptoms); chronic form (characterized by white or yellow streaks parallel to the main vein on leaves); acute form (presents attacks on xylem reducing sucrose production, and when in advanced stages can kill the plant) (Tokeshi and Rago, 1980). The leaf scald control is still scarce; it is only advisable to clean equipment and seedlings to reduce the propagation of the pathogen.
Considering the phytosanitary problems caused by Xanthomonas (X. citri and X. albilineans), it becomes necessary to research new forms of control, more efficient and less damaging to the environment.
Substances obtained from natural sources are important given the variability of such products. In general, natural molecules show greater structural diversity and several bioactive functions (Feher and Schmidt, 2003; Dayan et al., 2009; Newman and Cragg, 2016).
New microorganisms and natural products can be isolated from extreme environments, such as Antarctica (Pikuta et al., 2007). Many studies highlight the potential of the Antarctic environment in the production of bioactive metabolites (Furbino et al., 2014), antifungal substances (Svahn et al., 2015) and active compounds against Xanthomonas species, including X. citri (Moncheva et al., 2002; Encheva-Malinova et al., 2015; Puric et al., 2018; Vieira et al., 2018).
Thus, the present work is aimed to discover new fungi isolated from Antartic soils with potential for the production of bioactive compounds against X. citri and X. albilineans.
Strain of filamentous fungi
Sixty-one filamentous fungi were used in this study. Among them, twenty-three were isolated from soil under an iron bar in Deception Island (South Shetland Islands, Antarctica) (60º41'459''W), twenty-three from soil at 4.0ºC (62º05'092''S) and fifteen from soil at 1.6ºC (57º56'763''W), in King George Island (South Shetland Islands, Maritime Antarctica). All of filamentous fungi are being maintained by cryopreservation at -80ºC and Castellani at 4ºC at Central of Microbial Resources (CRM-UNESP).
Crude extracts production
The fungi were reactivated on agar malt broth petri dishes (2% malt and 1.5% agar) at 15ºC for 10 days. After their growth, disks of 5 mm of agar plus mycelium were transfered to 150 mL of 2% liquid malt broth in erlenmeyer flasks. This culture was shaken at 15ºC for 20 days at 150 rpm. After the period of incubation, the biomass and metabolic medium (supernatant) were separated using vacuum filtration.
Intracellular extracts
The biomass was extracted 3 times with 40 mL methanol by maceration.
Extracellular extracts
The supernatant was extracted 3 times with 60 mL ethyl acetate by liquid-liquid extraction. The extracts were concentrated in a low pressure rotary evaporator and dissolved in dimethylsulfoxide (DMSO) 10% (v/v) until the concentration of 30 mg/mL.
Cultivation of Xanthomonas
X. citri strain 306 (IBSBF 1594) was activated on agar NYG (3 g yeast extract, 5 g peptone and 20 mL glycerol) broth at 29ºC for 2 days. Then, bacterial cells were cultured in 30 mL of liquid NYG broth medium, shaken at 200 rpm and 29ºC until OD600 = 0.8.
X. albilineans (IBSBF 1387) was activated on agar YSG medium (5 g yeast extract, 5 g glucose, 0.5 g monobasic ammonium phosphate and 0.2 g anyhidrous dibasic potassium phosphate) broth at 29ºC for 5 days. Then, bacterial cells were cultured in 30 mL of liquid YSG broth medium, shaken at 200 rpm and 29ºC. The bacterial inoculum was standardized at 107 CFU/mL (OD600 = 0.9).
Anti-Xanthomonas activity assessment
The bioassay was accomplished in 96-well plates with triplicate of events and experiments, based in the Resazurin Microtiter Assay (REMA) method (Silva et al., 2013). For the bioassay against X. citri the maximum concentration tested of extracts was 2.1 mg/mL followed by microdilution until 0.016 mg/mL and for X. albilineans the maximum concentration tested of extracts was 3 mg/mL. Kanamycin (0.02 mg/mL) was used as positive control (PC), DMSO 1% (v/v) was used as vehicle control (VC) and negative control (NC) was made with medium (NYG) and inoculum. The inoculum was diluted (1:10) in NYG and aliquote of 10 µL was added to all wells used and standardized at 105 CFU/well. After a 16 h incubation at 29ºC 15 µL of resazurin 0.01% (w/v) was added and submitted to another incubation for about 1 h at the same temperature. The REMA was made in triplicate.
A Biotek Synergy H1MFD microplate reader was used to measure cell viability. It scans the fluorescence of the resorufin (resazurin reduced by microbial metabolism) at 530 nm (excitation) and 590 nm (emission). The fluorescence units (FU) detected by the reader were used to calculate growth inhibition in percentage, according to the formula:
Inhibition rate above 90% was considered as positive.
The minimal extract concentration needed for inhibition of 90% of cells (MIC90) was calculated based on the equation obtained from the regression curve constructed with the Rema bioassay data.
The bactericidal assay was performed as follows: A small aliquot from each well of the REMA plates was inoculated in petri dishes (150 x 15 mm, NYG medium) with 96-pin microplate replicator. After incubation for 48 h at 29ºC, it was possible to visually analyze the bacterial growth. The minimal concentration in which there was no bacterial growth was considered the minimal bactericidal concentration (MBC) of the extract.
Identification of the Antarctic fungi
DNA extraction was performed directly from the mycelium after the fungal growth in PDA medium for 7 to 14 days at 15°C, following a modified version of the CTAB method (Möller et al., 1992; Gerardo et al., 2004). ITS amplification was performed using primers ITS4 (5’TCCTCCGCTTATTGATATGC3’) and ITS5 (5’GGAAGTAAAAGTCGTAACAAGG3’) (Schoch et al., 2012).
PCRs were performed in a final volume of 25 μL (4 μL of dNTPs [1.25 mm each]; 5 μL of 5x buffer; 1 μL of BSA [1 mg/mL]; 2 μL of MgCl2 [25 mm]; 1 μL of each primer [10 μm]; 0.2 μL of Taq polymerase [5 U/μL], 2 μL of diluted genomic DNA [1:100] and 8.8 μL of sterile ultrapure water). PCR conditions for ITS were 94°C/3 min followed by 35 cycles at 94°C/1 min, 55°C/1 min and 72°C/2 min. Amplicons were purified using the Exonuclease I and Alkaline phosphatase (Thermo Scientific, Massachusetts, USA), according to the manufacturer's protocol. Samples were then quantified in NanoDrop® (Thermo Scientific) and sequenced using BigDye Terminator® v.3.1 kit (Applied Biosystems, California, USA), following the manufacturer’s instructions. Sequences obtained were compared with sequences deposited in the NCBI – GenBank database. Alignments were performed separately for each molecular marker using MAFFT v.7 (Katoh and Standley, 2013). The phylogenetic analyses were done using the program MEGA v. 7 (Tamura et al., 2013). The Kimura 2-parameter model (Kimura, 1980) was used to estimate evolutionary distance for the Neighbor-joining analysis. Branch support was calculated by bootstrap with 1,000 replicate runs.
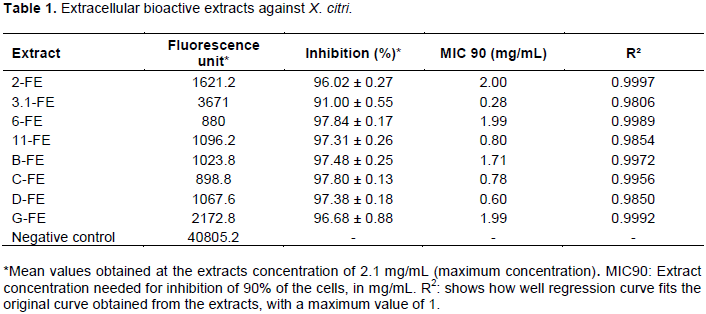
A total of 122 extracts was produced. All of them were tested against X. citri. Eight extracellular extracts presented, in the maximum concentration, averages of fluorescence measurements (Table 1) which when applied in the formula to calculate growth inhibition in percentage showed inhibition rates above 90% (Table 1) and were considered positive. The intracellular extracts showed no antibacterial activity against the tested Xanthomonas species.
The MIC90 values (Table 1) for the extracts of 2-Fe, 6-Fe, B-Fe and G-Fe were around 2.0 to 1.7 mg/mL, three extracts (11-Fe, C-Fe and D-Fe) presented values between 0.80 and 0.60 mg/mL, and the extract of 3.1-Fe attained the lowest MIC90 (0.28 mg/mL).
In the bactericidal assay, four extracts (2-Fe, 6-Fe, B-Fe and G-Fe) presented bacterial growth in the highest concentration tested (2.1 mg/mL), therefore they were considered bacteriostatic. The extracts of C-Fe, D-Fe, 3.1-Fe and 11-Fe were bactericidal, highlighting 3.1-Fe and 11-Fe, which had the lowest MBC value (1.0 mg/mL).
In the experiments with X. albilineans, two of the 122 extracts were considered positive, the extracts of 1.1-Fe and B-Fe presented a percentage of 97% inhibition of bacterial growth in the concentration 1.5 mg/mL. The MIC90 values for both extracts (1.1-Fe and B-Fe) were about 1.4 mg/mL. In the bactericidal bioassay the both extracts were bactericidal up to the concentration of 1.5 mg/mL (MBC).
In this study, eight bioactive fungi (1.1- Fe, 2-Fe, 3.1-Fe, 6-Fe, 11-Fe, B-Fe, C-Fe and G-Fe) were identified as belonging to the genus Pseudogymnoascus based on molecular taxonomy and phylogentic analysis (Figure 1).
The bioactive fungus D-Fe was identified as belonging to the genus Cladosporium (Figure 2). The taxonomic marker used (ITS) showed to be very conservative, since different species of the genus Cladosporium grouped in the same cluster of the Antarctic-derived fungus D-Fe.
Positive results of extracellular extracts may be associated with defense mechanism, since some microorganisms can produce substances that inhibit potential competitors (Smid and Lacroix, 2013; García et al., 2017). The samples tested are crude extracts, a reduction in MIC90 value is expected after purification and isolation of the bioactive molecules.
Based on this work, X. albilineans may be considered more resistant than X. citri, since only two extracts were inhibitory against X. albilineans. This also can be noted by the lower inhibition capacity of the antibiotic used as positive control (kanamicin) for this bacterium. In plantations, this resistance can be observed, since leaf scalding has no effective chemical combat and it is more difficult to contain than citrus canker.
The extract produced by the fungus B-Fe (Pseudogymnoascus) showed satisfactory bioactivity against both bacteria, and it can be considered as a promising microbial genetic resource in the fight against Xanthomonas.
The bioactive extracts (Table 1) were also tested, in another study, against X. euvesicatoria (bacterial spot on pepper and tomato) and X. axonopodis pv. manihotis (cassava bacteriosis) and only the fungus D-Fe showed activity against X. axonopodis pv. manihotis.
About identification, the Pseudogymnoascus ITS-rDNA sequences were highly similar, suggesting that it could be representatives of the same species. Amplicons were compared to homologous sequences deposited in the NCBI-GenBank database. The isolates showed 100% similarity with different strains of Pseudogymnoascus sp. Data from phylogenetic analysis showed that the isolates were placed in a separate cluster in relation to the other Pseudogymnoascus species (bootstrap value of 71%). These results suggest that Pseudogymnoascus strains could represent a new species. However, as reported by Chaturvedi et al. (2010), others taxonomic markers have
to be applied in order to get speciation for Pseudogymnoascus/Geomyces. Representatives of this genus are globally distributed and can develop in different conditions and extreme environment (Hayes, 2012), including Antarctic ecossystems (Arenz et al., 2006; Bridge and Spooner, 2012; Godinho et al., 2015). In the study reported by Wentzel et al. (2018) Pseudogymnoascus was the most abundant filamentous fungi found in antarctic samples of soil and marine sediments.
The isolates from Pseudogymnoascus have been reported as having activity against microorganisms (Furbino et al., 2014; Gonçalves et al., 2015). Figueroa et al. (2015) described four new nitroasterric acid derivatives with antibacterial and antifungal activity obtained from the cultures of a Pseudogymnoascus sp. fungus isolated from an Antarctic marine sponge. Other studies reported the bioactivity of this genus against Xanthomonas (Henríquez et al., 2014), including some from our research group, against X. citri, X. euvesicatoria and X. axonopodis pv. passiflorae (Vieira et al., 2018, Puric et al., 2018), but so far no molecules have been described.
Representatives of Cladosporium have also been reported in the Antarctic environments (Arenz et al., 2006; Bridge and Spooner, 2012; Wentzel et al., 2018). According to Tosi et al. (2002) Cladosporium is the most frequently observed genus in association with antarctic mosses. Antimicrobial molecules were reported as produced by representatives of this genus (Zhang et al., 2001; Xiong et al., 2009; Liang et al., 2018), but studies related to the bioactivity of Cladosporium against Xanthomonas are still scarce. In a previous study published by our research group, a representative of Cladosporium showed potencial to inhibit the cell growth of X. citri, X. euvesicatoria and X. axonopodis pv. passiflorae (Puric et al., 2018), corroborating the results achieved in the present study, where Cladosporium sp. (D-Fe) was able to produce bioactive molecules against X. citri.
The present study highlights the potential of bioprospecting of extrememophile microorganisms, in the search for new and more sustainable bioactive molecules, potentially able to substitute the use of copper in agriculture and in the combat of phytopathogens.
Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) (grants #2018/11747-3, #2015/20629-6 and #2016/07957-7) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (grant #401230/2016-3).
The authors declare that there is no conflict of interest.
REFERENCES
|
Arenz BE, Held BW, Jurgens JA, Farrell RL, Blanchette RA (2006). Fungal diversity in soils and historic wood from the Ross Sea Region of Antarctica. Soil Biology and Biochemistry 38(10):3057-3064.
Crossref
|
|
|
|
Bridge PD, Spooner BM (2012). Non-lichenized Antarctic fungi: transient visitors or members of a cryptic ecosystem? Fungal Ecology 5(4):381-394.
Crossref
|
|
|
|
|
Chaturvedi V, Springer DJ, Behr MJ, Ramani R, Li X, Peck MK, Ren P, Bopp DJ, Wood B, Samsonoff WA, Butchkoski CM, Hicks AC, Stone WB, Rudd RJ, Chaturvedi S (2010). Morphological and molecular characterizations of psychrophilic fungus Geomyces destructans from New York bats with white nose syndrome (WNS). PLoS One 5:e10783.
Crossref
|
|
|
|
|
Dayan FE, Cantrell CL, Duke SO (2009). Natural products in crop protection. Bioorganic and Medicinal Chemistry 17(12):4022-4034.
Crossref
|
|
|
|
|
Encheva-Malinova M, Vancheva T, Badzhinerov N, Koleva V, Tishkov S, Bogatzevska N, Moncheva P (2015). Antimicrobial activity of Antarctic streptomycetes against pepper bacterial spot causing agents. First National Conference of Biotechnology - Sofia 100:216-222.
|
|
|
|
|
Feher M, Schmidt JM (2003). Property Distributions: Differences between Drugs, Natural Products, and Molecules from Combinatorial Chemistry. Journal of Chemical Information and Computer Sciences 43(1):218-227.
Crossref
|
|
|
|
|
Figueroa L, Jiménez C, Rodríguez J, Areche C, Chávez R, Henríquez M, De la Cruz M, Díaz C, Segade Y, Vaca I (2015). 3‑Nitroasterric Acid Derivatives from an Antarctic Sponge-Derived Pseudogymnoascus sp. Fungus. Journal of Natural Products 78:919-923.
Crossref
|
|
|
|
|
Furbino LE, Godinho VM, Santiago IF, Pellizari FM, Alves TMA, Zani CL, Junior PAS, Romanha AJ, Carvalho AGO, Gil LHVG, Rosa CA, Minnis AM, Rosa LH (2014). Diversity Patterns, Ecology and Biological Activities of Fungal Communities Associated with the Endemic Macroalgae Across the Antarctic Peninsula. Microbial Ecology 67:775-787.
Crossref
|
|
|
|
|
García C, Rendueles M, Díaz M (2017). Microbial amensalism in Lactobacillus casei and Pseudomonas taetrolens mixed culture. Bioprocess and Biosystems Engineering 40(7):1111-1122.
Crossref
|
|
|
|
|
Gerardo NM, Mueller UG, Price SL, Currie CR (2004). Exploiting a mutualism: parasite specialization on cultivars within the fungus-growing ant symbiosis. Proceedings of the Royal Society Biological Sciences 271:1791-1798.
Crossref
|
|
|
|
|
Gonçalves VN, Carvalho CR, Johann S, Mendes G, Alves TMA, Zani CL, Junior PAS, Murta SMF, Romanha AJ, Cantrell CL, Rosa CA, Rosa LH (2015). Antibacterial, antifungal and antiprotozoal activities of fungal communities present in different substrates from Antarctica. Polar Biology 38(8):1143-1152.
Crossref
|
|
|
|
|
Godinho VM, Gonçalves VN, Santiago IF, Figueredo HM, Vitoreli GA, Schaefer CEGR, BArbosa EC, Oliveira JG, Alves TMA, Zani CL, Junior PAS, Murta SMF, Romanha AJ, Kroon EG, Cantrell CL, Wedge DE, Duke SO, Ali A, Rosa CA, Rosa LH (2015). Diversity and bioprospection of fungal community present in oligotrophic soil of continental Antarctica. Extremophiles 19(3):585-596.
Crossref
|
|
|
|
|
Hayes MA (2012). The Geomyces fungi: ecology and distribution. Bioscience 62:819-823.
Crossref
|
|
|
|
|
Henríquez M, Vergara K, Norambuena J, Beiza A, Maza F, Ubilla P, Araya I, Chávez R, San-Martín A, Darias J, Darias MJ, Vaca I (2014). Diversity of cultivable fungi associated with Antarctic marine sponges and screening for their antimicrobial, antitumoral and antioxidant potential. World Journal of Microbiology and Biotechnology, pp.30-65.
Crossref
|
|
|
|
|
Katoh K, Standley DM (2013). MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Molecular Biology and Evolution 30:772-780.
Crossref
|
|
|
|
|
Kimura M (1980). A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. Journal of Molecular Evolution 16:111-120.
Crossref
|
|
|
|
|
Leyns F, De Cleene M, Swings JG, Ley JD (1984). The host range of the genus Xanthomonas. The Botanical Review 50(3):308-356.
Crossref
|
|
|
|
|
Liang X, Huang ZH, Ma X, Qi SH (2018). Unstable Tetramic Acid Derivatives from the Deep-Sea-Derived Fungus Cladosporium sphaerospermum EIODSF 008. Marine Drugs 16(11):448.
Crossref
|
|
|
|
|
Möller EM, Bahnweg G, Sandermann H, Geiger HH (1992). A simple and efficient protocol for isolation of high molecular weight DNA from filamentous fungi, fruit bodies, and infected plant tissues. Nucleic Acids Research 20:6115-6116. PMID: 1461751
Crossref
|
|
|
|
|
Bogatzevska N (2002). Characteristics of soil actinomycetes from Antarctica. Journal of Culture Collections 3:3-14.
|
|
|
|
|
Newman DJ, Cragg GM, (2016). Natural products as sources of new drugs from 1981 to 2014. Journal of Natural Products 79(3):629-661.
Crossref
|
|
|
|
|
Pikuta EV, Hoover RB, Tang J (2007). Microbial Extremophiles at the Limits of Life. Critical Reviews in Microbiology 33:183-209.
Crossref
|
|
|
|
|
Puric J, Vieira G, Cavalca LB, Sette LD, Ferreira H, Vieira MLC, Sass DC (2018). Activity of antarctic fungi extracts against phytopathogenic bacteria. Letters in Applied Microbiology 66:530-536.
Crossref
|
|
|
|
|
Rocha CHB, Azevedo LP (2017). Avaliação da presença de metais pesados nas águas superficiais da Bacia do Córrego São Mateus, Juiz de Fora (MG), Brasil. Revista Espinhaço - UFVJM 33-44. ISSN 2317-0611
|
|
|
|
|
Ryan RP, Vorhölter FJ, Potnis N, Jones BJ, Sluys MAV, Bogdanove AJ, Dow JM (2011). Pathogenomics of Xanthomonas: understanding bacterium-plant interactions. Nature Reviews Microbiology 9:344-355.
Crossref
|
|
|
|
|
Schaad NW, Postnikova E, Lacy G, Sechler A, Agarkova IV, Stromberg PE, Stromberg VK, Vidaver AK (2006). Emended classification of xanthomonad pathogens on citrus. Papers in Plant Pathology, p. 96.
Crossref
|
|
|
|
|
Schoch CL, Seifert KA, Huhndorf S, Robert V, Spouge JL, Levesque CA, Chen W (2012). Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. The Fungal Barcoding Consortium. Proceedings of the National Academy of Sciences of the United States of America 109:6241-6246.
Crossref
|
|
|
|
|
Silva IC, Regasini LO, Petrônio MS, Silva DHS, Bolzani VS, Belasque Junior J, Sacramento LVS, Ferreira H (2013). Antibacterial activity of alkyl gallates against Xanthomonas citri subsp. citri. Journal of Bacteriology 195(1):85-94.
Crossref
|
|
|
|
|
Smid EJ, Lacroix C (2013). Microbe-microbe interactions in mixed culture food fermentations. Current Opinion in Biotechnology 24:148-154.
Crossref
|
|
|
|
|
Svahn KS, Chryssanthou E, Olsen B, Bohlin L, Göransson U (2015). Penicillium nalgiovense Laxa isolated from Antarctica is a new source of the antifungal metabolite amphotericin B. Fungal Biology and Biotechnology 2:1-8.
Crossref
|
|
|
|
|
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013). MEGA6: molecular evolutionary genetics analysis version 6.0. Molecular Biology and Evolution 30:2725-2729.
Crossref
|
|
|
|
|
Tessmann C (2002). Caracterização molecular de Xanthomonas campestris pv pruni pela técnica de RAPD e relação com a planta hospedeira e com a produção. viscosidade e composição química da xantana. 33p
|
|
|
|
|
Tokeshi H, Rago A (1980). Doenças da cana-de-açúcar. In: Galli F, Tokeshi H, Carvalho PT, et al. (1980). Manual de Fitopatologia: doenças das plantas e seu controle (Vol. 4). Biblioteca Agronômica Ceres.
|
|
|
|
|
Tosi S, Casado B, Gerdol R, Caretta G (2002). Fungi isolated from Antarctic mosses. Polar Biology 25(4):262-268.
|
|
|
|
|
Vieira G, Puric J, Morão LG, Santos JA, Inforsato FJ, Sette LD, Ferreira H, Sass DC (2018). Terrestrial and marine Antarctic fungi extracts active against Xanthomonas citri subsp. citri. Letters Applied Microbiology 67:64-71.
Crossref
|
|
|
|
|
Wentzel LCP, Inforsato FJ, Montoya QV, Rossin BG, Nascimento NR, Rodrigues A, Sette LD (2018). Fungi from Admiralty Bay (King George Island, Antarctica) Soils and Marine Sediments. Microbial Ecology 1-13.
Crossref
|
|
|
|
|
Xiong H, Qi S, Xu Y, Miao L, Quian PY (2009). Antibiotic and antifouling compound production by the marine-derived fungus Cladosporium sp. F14. Journal of Hydro-environment Research 2(4):264-270.
Crossref
|
|
|
|
|
Zhang H, Tomodai H, Tabata N, Miura H, Namikoshi M, Yamaguchi Y, Masuma R, Omura S (2001). Cladospolide D, a new 12-membered macrolide antibiotic produced by Cladosporium sp. FT-0012. The Journal of Antibiotics 54(8):635-641.
Crossref
|
|