ABSTRACT
In this study, a total of 9 morphologically and biochemically different yeast species were isolated from Nigerian fermented food product, ‘ogi’. These yeast species were screened for probiotic properties such as antibacterial activities, acid and bile tolerance. Six species out of the yeast isolates that survived pH 2 were grown in 0.3% bile salts which were monitored by spectrophotometer for 5 h. Three yeast species with the highest growth performance measured by increment in optical density were identified as Pichia kudriavzevii by sequencing the conserved (D1/D2) region of 16S rDNA. Among the three selected strains, isolate MH458240 (M9) showed the highest acid tolerance of 5.80±0.20 log cfu/ml while isolate MH458239 (M5) manifested the least acid tolerance of 3.80±0.20 log cfu/ml. At 0.3% bile salts concentration, isolate MH458239 (M5) demonstrated the highest growth rate of 1.110±011. The antibacterial activities of the three selected yeast strains were also determined. While isolates MH458240 (M9) and MH458239 (M5) inhibited the growth of the tested pathogens, isolate MH458238 (M4) could not inhibit the growth of any of the pathogens used. Out of the three strains of P. kudriavzevii used, isolate MH458240 (M9) which demonstrated the best overall probiotic potential was found to contain only three open reading frames (ORF) while the two other isolates contained five. Isolate MH458240 (M9) was found to be in a complete different cluster from the two other strains.
Key words: Yeasts, fermented food, Pichia kudriavzevii, probiotic properties, open reading frames (ORF).
‘Ogi’, a fermented cereal gruel considered as a beverage and also a weaning food in Nigeria, could be processed from maize, although sorghum or millet can also be employed as the substrate for fermentation. Fermentation is one of the oldest technologies used for food processing as well as preservation. It has been found to enhance food flavor, produce useful enzymes, vitamins, amino acids and anti-microbial products (Vlasova et al., 2016). Integral parts of fermentation process are non-pathogenic, useful microorganisms commonly referred to as probiotics (Corgan et al., 2007; Kalui et al., 2008). Researchers in the field of science have changed their
view of the role of food as being just an energy giver and body builder to having an important role of possessing active substances/ingredients that affect the health of consumers positively. Such foods that are beneficial to health are referred to as functional foods since they go beyond keeping the consumers’ health to reducing the risk of chronic diseases (Lei et al., 2008; Vasiljevic and shah, 2008).
The two major types of microorganisms associated with traditional fermented foods and beverages are yeasts and bacteria. Yeasts are found in different classes of foods ranging from plant to animal origins where they have a notable impact on food safety and organoleptic properties (FAO and WHO, 2001; Chelliah et al., 2016). Various studies have reported the occurrence of different yeast genera such as Saccharomyces, Candida, Debaryomyces, Kluyveromyces, and Pichia species, in African traditional fermented food (Kumura et al., 2004; Mahasneh and Abbas, 2010).
Probiotics are live microorganisms, which when administered in adequate amount confer a health benefit on the host (Sanchez et al., 2017). This definition suggests that there must be a measurable physiologic benefit to the host who used the probiotic product (Vandenplas et al., 2014). A joint FAO/WHO working group in 2002 released guidelines for the evaluation of probiotics in food. The basic requirements needed include assessment of strain identity, resistance to gastrointestinal acidity, digestive enzymes, bile acids, antimicrobial activity against potentially pathogenic bacteria and in vivo studies for substantial health effects in the target host. However, the ultimate criterion to select a successful probiotic strain is the ability to confer to consumers a health benefit (Åukaszewicz, 2012). The commonly used probiotics include Lactic Acid Bacteria especially those of the genera Lactobacillus, Bifidobacteria and yeast of the genus Saccharomyces (Vlasova et al., 2016). Recently, it was reported that all probiotics are not equally effective (Szajewska et al., 2016). Both in vitro and in vivo experiments suggested that probiotic efficiency differs from strain to strain among tested probiotic strains. Different strains of potential probiotics have different mechanism of action and the ability to elicit immune response (McFarland, 2014; Goldstein et al., 2015).
Many characteristic attributes distinguished yeasts as probiotic such as their robust size, ability to tolerate low pH, oxygen, water activity, high osmotic pressure, nutritional flexibility, enzymatic activity and ability to produce several useful metabolites (Fredlund et al., 2002; Lukaszewics, 2012). However, very limited numbers of yeasts are considered as probiotics with proven clinical efficiency in double blind studies (Foligne et al., 2010; Riaz et al., 2012; Lohith and Appaiah, 2014).
Pichia kudriavzevii (the teleomorph of Candida krusei) is one of the microflora predominant in the fermentation of maize for ‘ogi’ production. It has also been implicated in the fermentation of fruit juices and cocoa beans where it has been found helpful in amplifying the aroma of chocolate (Oberoi et al., 2012). P. kudriavzevii can be found in the soil as well as on the surfaces of fruits and vegetables (Chan, 2012). Although this organism was present on plant produce, it was not considered as a species that causes food spoilage (Fleet, 2011).
P. kudriavzevii has the potential of producing toxin that is able to kill several pathogenic microorganisms thereby contributing to food preservation (Bajaj et al., 2013). This organism has also been considered a potential probiotics for its ability to assimilate cholesterol (Psomas et al., 2003). In this study therefore, the probiotic potentials of three strains of P. kudriavzevii isolated from a Nigerian traditional fermented food were evaluated.
Sample collection
Samples of ogi were randomly collected from retailers within Ogbomoso Township, Oyo State, Nigeria. All samples were collected in sterile, glass screw-capped bottles and transported immediately to the laboratory for analysis.
Isolation of yeasts from collected samples
Ten-fold serial dilutions (10-1 to 10-9) of the samples were made using sterile peptone water. 1 mL aliquot of 10-4, 10-6 and 10-8 dilutions were pour plated in duplicates on yeast peptone dextrose (YPD) agar (per Litre; 10 g yeast extract, 20 g peptone, 20 g glucose and 20 g agar) (LAB M, UK) supplemented with chloramphenicol (100 mg/l) and incubated at 25°C for 48 h.
Colonies with distinct morphological differences such as colour, shape and size were streaked out as representative colony and were then purified by subculturing on Yeast Extract Peptone (YPD) agar to obtain pure culture of the isolates. The purified isolates were maintained on YPD agar slants at 4°C until required for identification (Kurtzman et al., 2011).
Screening for probiotic properties
Tolerance to pH 2.0
The isolates were subjected to a pH 2.0 assay in order to select the resistant isolates for further studies. Yeasts cells were cultivated in YPD broth for 24 h. The culture was centrifuged (4000 g (rpm) for 10 min at 24°C) and washed two times in 0.1% w/v peptone water pH 7.0. The cell cultures (optical density of 0.2 at 600 nm) in peptone water corresponding to approximately 108 cell/ml were centrifuged and resuspended in YPD broth with pH adjusted to 2.0 using 1 N HCl were incubated for 3 h at 37°C. Samples (10 µL) were obtained at time 0 and at 3 h and inoculated in YPD agar plates. Duplicate plates were obtained and tolerance to pH 2.0 was indicated by subsequent growth on agar plates after 48 h of incubation at 37°C as described by Ramos et al. (2013).
Determination of bile tolerance
The method described by Guo et al. (2009) was modified and used to determine the effect of bile on the growth rate of acid tolerant isolates. Growth performance was expressed as the difference in optical density at 600 nm between the initial and 5 h of incubation 0.3% bile salts concentration (SIGMA ALDRICH, Co, St Louis, USA). The experiment was performed in duplicate.
Molecular identification of yeast isolates
Three acid and bile tolerant yeast species were selected for molecular identification. Genomic DNA of the yeast isolates were extracted using phenol-chloroform extraction method (Green and Sambrook 2017). The D1/D2 region of large subunit of 26S rDNA of selected yeast species was amplified with universal primers, 5’ -GCATATCAA TAAGCGGAGGAAAAG-3’ and 5’ -GGTCCGTGTTTCAA GACGG-3’ for forward and reverse, respectively (Kurtzman and Robnett, 1998) in a thermocycler (MJ RESEARCH, INC, programmable thermal controller, BIO-RAD). The amplicon was sequenced using the Sanger’s sequencing method (Beckman coulter, USA) and the nucleotide sequences were aligned in genbank using the BLAST program (Website: www.ncbi.nlm.nih.gov/blast). Phylogenetic tree was constructed using MEGA 4.1 software programme (Tamura et al., 2007).
Comparative genomic and phylogenetic analysis
The comparative genomic analysis was carried out on the nucleotide sequences, the exploitation of evolutionary information to draw powerful and robust inferences about phylogenetic history using webPRANK (https://www.ebi.ac.uk/goldman-srv/webprank/). The phylogenetic analysis was carried out through multiple sequence alignment of the nucleotide sequences using MEGA 7.0. Approximately-maximum-likelihood tree was created with MEGA (version 7.0).
Antibacterial activity against enteric pathogens
The double layer method of Maia et al. (2001) was adopted in the determination of the inhibitory effects of yeast isolates against some enteric pathogens. The yeasts were tested against Escherichia coli, Staphylococcus aureus, Pseudomonas species, Klebsiella species and Proteus species. A 24 h old culture of yeast isolates were prepared in YPD broth and was individually inoculated onto YPD agar plates by swabbing an inch by 1.5 inch area in the centre of each plate. The plates were incubated at 37°C for 48 h. These plates were overlaid individually with 10 mL of molten nutrient agar that has been previously inoculated with 1 mL of the pathogen culture. The agar was allowed to solidify and the plates were incubated aerobically at 37°C for 24 h. Plates were examined for growth of the yeast strains with their ability to inhibit the growth of the pathogens.
Statistical analysis
Results were expressed as mean+/- standard error. Multiple comparisons were performed by one-way analysis of variance (ANOVA) followed by least significance difference (LSD) test and statistical significance was accepted at values of P<0.05 using SPSS.
Tolerance to pH 2.0
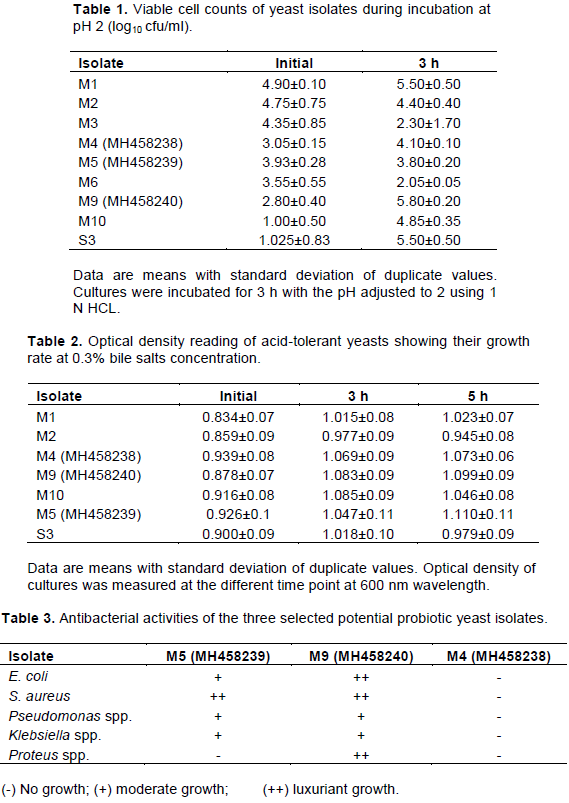
Ability of the yeast isolates to tolerate acidic pH is shown in Table 1. The highest survival rate was observed in isolate MH458240 (M9) with initial count of 2.80±0.40 and 5.80±0.20 cfu/ml after 3 h of incubation. The least survival was observed in isolate M6 with initial count of 3.55±0.55 cfu/ml and declined after 3 h of incubation to 2.05±0.05 cfu/ml. A slight decline was observed in isolate MH458239 (M5) from initial viable count of 3.93±0.28 to 3.80±0.20. Isolates M3 and M2 also declined significantly (P<0.05) after 3 h of incubation, which suggested that they could not be successful candidates for probiotic use.
Bile tolerance test
Without the addition of bile salt an overall increase was observed in the rate of growth at 5 h of incubation (result not shown). At 0.3% bile salt concentration, isolate MH458239 (M5) manifested the highest significant increase in its rate of growth with absorbance of 0.926±0.1, 1.047±0.11 and 1.11±0.11 at 0, 3 and 5 h of incubation, respectively (Table 2). Isolates MH458238 (M4) and MH458240 (M9) also showed significant high growth rate with initial absorbance reading of 0.939±0.08 and 0.878±0.07, respectively which increased to 1.073±0.06 and 1.099±0.09 at the end of 5 h of incubation.
Antibacterial activity
The result of the antagonistic effect of the three strains of selected yeasts against E. coli, Pseudomonas spp., S. aureus, Klebsiella spp. and Proteus spp. is shown in Table 3. Yeast isolates MH458239 (M5) and MH458240 (M9) inhibited the growth of the test pathogens which was indicated by outgrowth of the yeast culture, although isolate MH458239 (M5) could not outgrow Proteus species. Isolate MH458238 (M4) did not express any antagonistic effect against any of the bacterial enteric pathogens and therefore, no growth of the yeast was observed on the plate.
Comparative genomic and phylogenetic analysis
The comparative genomics of the three strains of P. kudriavzevii isolates obtained is shown in Table 4. This result was generated using webPRANK for the sequence alignment of the three strain nucleotide sequences. From the alignment of the sequences, isolate MH458240 (M9) had several missing gaps when compared with the other two isolates.
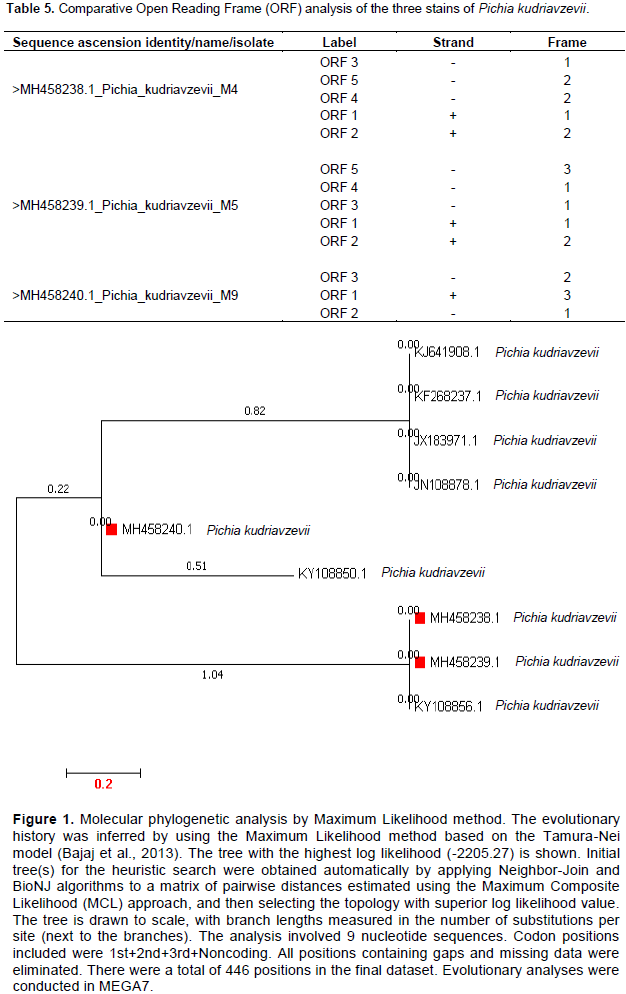
The various open reading frame (ORF) positions in the nucleotide sequences of the selected three strains are shown in Table 5. While isolate MH458240 (M9) had three ORF; isolates MH458238 (M4) and MH45839 (M5) each had five. From the phylogeny shown in Figure 1, it was observed that isolates MH458238 (M4) and MH458239 (M5) were more closely related as shown in the same cluster while isolate MH458240 (M9) was in a complete different cluster.
Passage through the stomach is the first line of barrier to be overcome by probiotic organisms when administered through the oral route. In this present study, yeasts were isolated from a Nigerian traditional fermented food, ogi. It was observed that majority of the tested yeast isolates could thrive at pH 2, the average pH of the gastric juice. After about 3 h of ingestion, the stomach would empty its contents into the intestine where some digestive enzymes act on it. One of those important enzymes is the bile salts. A successful potential probiotic candidate should be able to grow and replicate significantly in the presence of bile salts so that it can produce its metabolites and deliver the health benefit for which it was administered. It was observed that some of the yeast isolates also grew well in medium supplemented with bile salts.
The best acid and bile tolerant yeast strains were selected for molecular identification. They were isolates MH458240 (M9), MH458239 (M5) and MH458238 (M4) which were all identified as P. kudriavzevii. Several works have indicated that P. kurdriavzevii tolerate acid and bile well. For example, Greppi et al. (2016) showed that out of the 93 yeast strains screened for their tolerance to pH 2 and 0.3% bile salts, P. kudriavzevii isolates gave the best results in thriving well in these culture conditions.
Ogunremi et al. (2015) also isolated P. kudriavzevii from ogi, a Nigerian traditional fermented food. P. kudriavzevii has been implicated in all sorts of traditionally fermented food and beverages. P. kurdriavzevii was isolated from fermented African cassava, Ghanaian fermented cocoa bean, tea and ginger tea (Kurtzman et al., 2011; Chan et al., 2012). van Rijswijck et al. (2017) also isolated P. kudriavzevii from fermented Masau fruits in Zimbabwe. The yeast has also been isolated from non-fermented food origin like chicken egg, human heart blood, swine waste, sputum and human feaeces and was considered as potential producer of bioethanol and phytase (Chan et al., 2012). Ogunremi et al. (2015) used this yeast as a starter culture for the development of cereal-based functional food.
From the three strains of P. kurdriavzevii used in this work, isolate MH458240 (M9) showed the highest acid tolerance of 5.80±0.20 log cfu/ml while isolate MH48239 (M5) manifested the least acid tolerance of 3.80±0.20 log cfu/ml. At 0.3% bile salts concentration, isolate MH48239 (M5) demonstrated the highest growth rate of 1.110±011 while isolate MH48238 (M4) showed the least bile tolerance. While both isolates MH458240 (M9) and MH458239 (M5) inhibited the growth of the tested pathogens, isolate MH458238 (M4) could not inhibit the growth of any of the pathogens used. These differences in the probiotic potentials of the three strains of P. kurdriavzevii could probably be due to the three ORF found in isolate MH458240 (M9) compared to five ORF in isolates MH458238 (M4) and MH458239 (M5). These differences in the ORF number may also be the reason why isolate MH458240 (M9) was in a completely different cluster from the two other isolates.
It can be concluded from this work that probiotic properties are strain specific as demonstrated by these three strains of P. kudriavzevii.
The authors have not declared any conflict of interests.
REFERENCES
|
Bajaj B, Raina S, Signh S (2013). Killer Toxin from a Novel Killer Yeast Pichia kudriavzevii RY55 with Idiosyncratic Antibacterial Activity. Journal of Basic Microbiology 53:645-656.
Crossref
|
|
|
|
Chan GF, Gan HM, Ling HL, Rashid NA (2012). Genome sequence of Pichia kudriavzevii M12, a potential producer of bioethanol and phytase. Eukaryot Cell 11:10.
Crossref
|
|
|
|
|
Chelliah R, Ramakrishnan SR, Prabhu, PR, Antony U. (2016). Evaluation of antimicrobial activity and probiotic properties of wild-strain Pichia kudriavzevii isolated from frozen idlibatter. Yeast 33(8):385-401.
Crossref
|
|
|
|
|
Corgan TM, Bresford TP, Steele J, Broadbent J, Shah NP, Ustunol Z (2007). Advances in starter cultures. Journal of Dairy Science 90:4005-4021.
Crossref
|
|
|
|
|
Fleet G (2011). The Yeasts. Elsevier IBSN: 978-0-444-52149-1
|
|
|
|
|
Foligne B, Dewulf J, Vandekerckove P, Pignede G, Pot B (2010). Probiotic yeasts: Anti-inflammatory potential of various non-pathogenic strains in experimental colitis in mice. World Journal of Gastroenterology 16(17):2134-2145.
Crossref
|
|
|
|
|
Food and Agriculture Organization (FAO), World Health Organization (WHO) (2001). Report of a joint FAO/WHO expert consultation on evaluation of health and nutritional properties of probiotics in food including powder milk with live lactic acid bacteria. 1-4 October 2001, Córdoba, Argentina.
|
|
|
|
|
Food and Agriculture Organization (FAO), World Health Organization (WHO) (2002). Report of a Joint FAO/WHO working group on drafting guidelines for the evaluation of probiotics in food.
|
|
|
|
|
Fredlund E, Druvefors U, Boysen EM, Lingsten K, Schnurer J (2002). Physiological characteristics of the biocontrol yeast Pichia anomala J121. FEMS Yeast Research 2:395-402.
Crossref
|
|
|
|
|
Goldstein EJ, Tyrrell KL, Citron DM (2015). Lactobacillus species: taxonomic complexity and controversial susceptibilities. Clinical Infectious Diseases 60(Suppl 2):S98-107.
Crossref
|
|
|
|
|
Green MR, Sambrook J (2017). Isolation of high-molecular-weight DNA using organic solvents. Cold Spring Harb Protocols 4:pdb-rot093450.
Crossref
|
|
|
|
|
Greppi A, Saubade F, Botta C, Humblot C, Guyot JP, Cocolin L (2016). Potential probiotic Pichia kudriavzevii strains and their ability to enhance folate content of traditional cereal-based African fermented Food and Food Microbiology 62:169-77.
Crossref
|
|
|
|
|
Guo Z, Wang J, Yan L, Chen W, Liu X, Zhang H (2009). In vitro comparison of probiotic properties of Lactobacillus casei Zhang, a potential new probiotic, with selected probiotic strains LWT. Food Science and Technology 42:1640-1646.
Crossref
|
|
|
|
|
Kalui CM, Mathara JM, Kutima PM, Kiiyukia C, Wongo LE (2008). Partial characterization and identification of lactic acid bacteria involved in the production of ikii: a traditional fermented maize porridge by the Kamba of Kenya. Journal of Tropical Microbiology and Biotechnology 4(1):3-15.
Crossref
|
|
|
|
|
Kumura H, Tanoue Y, Tsukahara M, Tanaka T, Shimazaki K (2004). Screening of dairy yeast strains for probiotic applications. Journal of Dairy Science 87(12):4050-4056.
Crossref
|
|
|
|
|
Kurtzman CP, Robnett, CJ (1998). Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie van Leeuwenhoek 73(4):331- 371.
Crossref
|
|
|
|
|
Kurtzman CP, Fell JW, Boekhout T (2011). The Yeasts, a Taxonomic Study. Volume1. Fifth edition. Elsevier.
|
|
|
|
|
Lei V, Friis H, Michaelsen KF (2008). Spontaneously fermented millet product as a natural probiotic treatment for diarrhoea in young children: An intervention study in Northern Ghana. International Journal of Food Microbiology 110:246-253.
Crossref
|
|
|
|
|
Lohith K, Anu Appaiah KA (2014). In vitro probiotic characterization of yeasts of food and environmental origin. International Journal of Probiotics and Prebiotics 9(3):87-92.
|
|
|
|
|
Åukaszewicz M (2012). Saccharomyces cerevisiae var. boulardii - Probiotic Yeast. Intech, 557 Chapter 16.
Crossref
|
|
|
|
|
Mahasneh AM, Abbas MM (2010). Probiotics and Traditional Fermented Foods: The Eternal Connection. Jordan Journal of Biological Sciences 3(4):133-140.
|
|
|
|
|
Maia OB, Duarte R, Silva AM (2001). Evaluation of the components of a commercial probiotic in gnotobiotic mice experimentally challenged with Salmonella enterica subsp. enterica ser. Typhimurium. Veterinary Microbiology 79(2):183-189.
Crossref
|
|
|
|
|
McFarland LV (2014). Use of probiotics to correct dysbiosis of normal microbiota following disease or disruptive events: a systematic review. BMJ Open 4(8).
Crossref
|
|
|
|
|
Oberoi H, Babbar N, Sandhu S, Dhaliwal S, Kaur U, Chadha B, and Bhargav V (2012). Ethanol production from alkali-treated rice straw via simultaneous saccharification and fermentation using newly isolated thermotolerant Pichia kudriavzevii HOP-1. Journal of Industrial Microbiology and Biotechnology 39(4):557-556.
Crossref
|
|
|
|
|
Ogunremi OR, Sanni AI, Agrawal R (2015). Probiotic potentials of yeasts isolated from some cereal-based Nigerian traditional fermented food products. Journal of Applied Microbiology 119:797-808.
Crossref
|
|
|
|
|
Psomas EI, Fletouris DJ, Litopoulou-Tzanetaki E, Tzanetakis N (2003). Assimilation of cholesterol by yeast strains isolated from infant feces and Feta cheese. Journal of Dairy Science 86:3416-3422.
Crossref
|
|
|
|
|
Ramos CL, Thorsen L, Schwan RF, Jespersen L (2013). Strain-specific probiotics properties of Lactobacillus fermentum, Lactobacillus plantarum and Lactobacillus brevis isolates from Brazilian food products. Food Microbiology 36(1):22-29.
Crossref
|
|
|
|
|
Riaz M, Alam S, Malik A, Ali SM (2012). Efficacy and safety of Saccharomyces boulardii in acute childhood diarrhea: A double blind randomised controlled trial. Indian Journal of Pediatrics 79(4):478-482.
Crossref
|
|
|
|
|
Sanchez B, Delqado S, Blanco-Miquez A, Lourenco A, Gueimonde M, Marqolles A (2017). Probiotics, gut microbiota and their influence on host health and disease. Molecular Nutrition and Food Research 61:1.
Crossref
|
|
|
|
|
Szajewska H, Canani RB, Guarino A, Hojsak I, Indrio F, Kolacek S (2016). Probiotics for the prevention of antibiotic-associated diarrhea in children. Journal of Pediatric Gastroenterology and Nutrition 62(3):495-506.
Crossref
|
|
|
|
|
Tamura K, Dudley J, Nei M, Kumar S (2007). MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Molecular Biology and Evolution 24:1596-1599.
Crossref
|
|
|
|
|
van Rijswijck IMH, Derks MFL, Abee T, De Ridder D, Smid EJ (2017). Genome sequences of Cyberlindnera fabianii 65, Pichia kudriavzevii 129 and Saccharomyces cerevisiae 131 isolated from fermented masau fruits in Zimbabwe. Genome Announc 5:e 00064-17
Crossref
|
|
|
|
|
Vandenplas Y, Huys G, Daube G (2014). Probiotics: an update. Jornal de Pediatria.
Crossref
|
|
|
|
|
Vasiljevic T, Shah NP (2008). Probiotics. From Metchnikoff to bioactives. International Journal of Dairy 18:714-728.
Crossref
|
|
|
|
|
Vlasova AN, Kandasamy S, Chattha KS, Rajashekara G, Saif LJ (2016). Comparison of probiotic Lactobacilliand Bifidobacteriaeffects, immune responses and rotavirus vaccines and infection in different host species. Veterinary Immunology and Immunopathology 172:72-84.
Crossref
|
|