Full Length Research Paper

ABSTRACT
Moisture deficit stress, one of the abiotic stresses, affects sugarcane growth and development and reduces cane and sugar yields. Transcription Factors (TFs) are master regulatory proteins in all living cells which have the capability of activating or repressing transcription of stress responsive genes in order to activate the stress tolerance mechanism. Study of expression profiles of TF genes which regulate the expression of stress responsive genes help to elucidate the regulatory biology of stress tolerance. Expression of 17 sugarcane TF genes in moisture deficit stress sensitive and tolerant varieties under different moisture deficit stress conditions were quantified in quantitative real-time PCR. Expression of seven TF genes namely, WRKY, NAC, bZIP, DREB, G2 like, Homeobox and TUB showed significant difference between the stress tolerant and susceptible varieties under both moderate and severe moisture deficit stress conditions. In stress tolerant variety, of these seven TF genes, bZIP showed highest expression both under moderate (22.39 fold) and severe stress (13.45 fold) conditions than other TF genes. Expression of bZIP gene in moisture deficit stress susceptible variety was significantly low under moderate (1.09 fold) and severe (3.63 fold) moisture deficit stress condition. GRAS TF gene under moderate stress condition (4 fold) and Homeobox gene under severe stress condition (6.06 fold) showed highest expressions than other TF genes in moisture stress susceptible variety. These differentially expressed TFs among the moisture stress tolerant and sensitive varieties hold promise for improving abiotic stress tolerance in sugarcane through their use as the potential candidate genes in marker assisted selection and in genetic transformation.
Key words: Transcription factors, moisture deficit stress, qRT- PCR, sugarcane.
INTRODUCTION
MATERIALS AND METHODS
RESULTS AND DISCUSSION
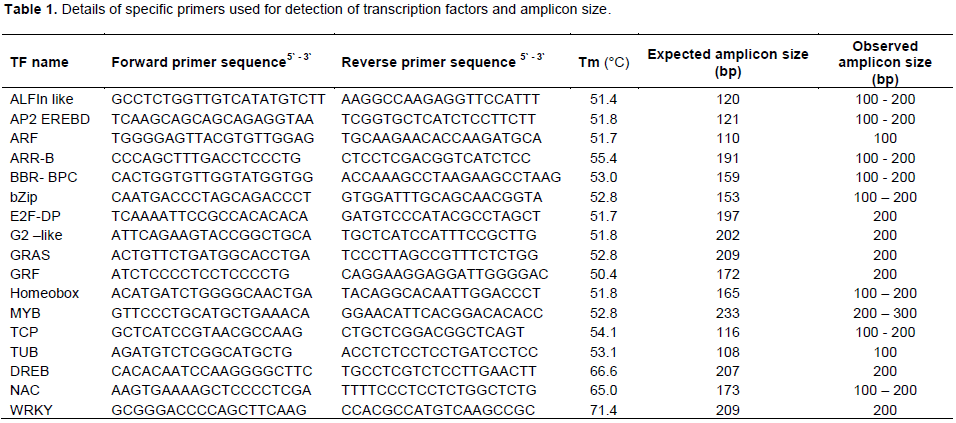
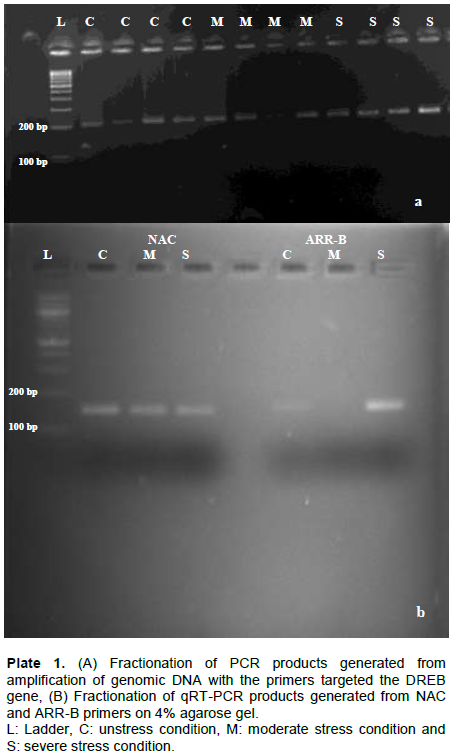
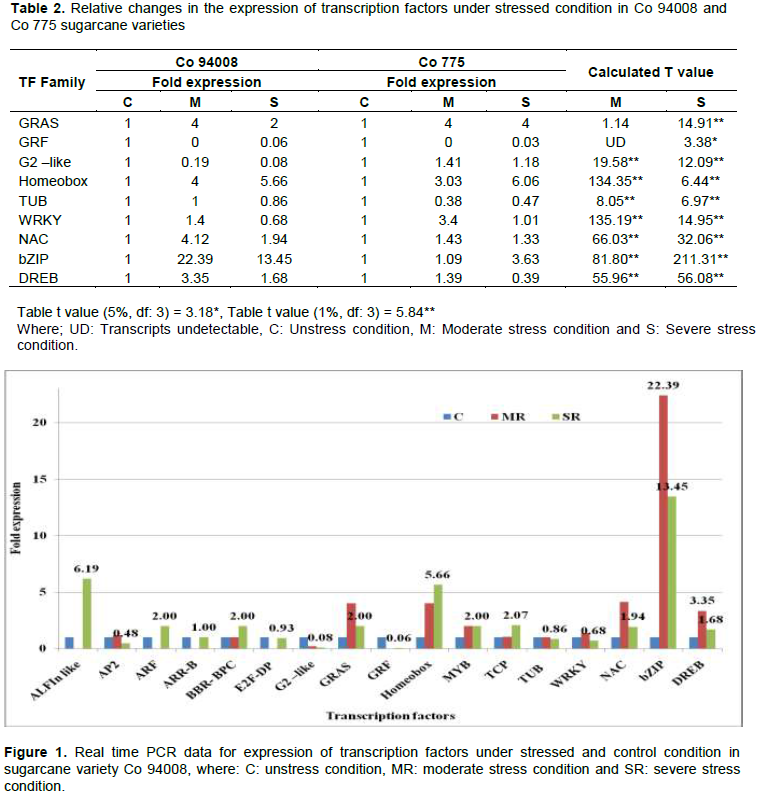
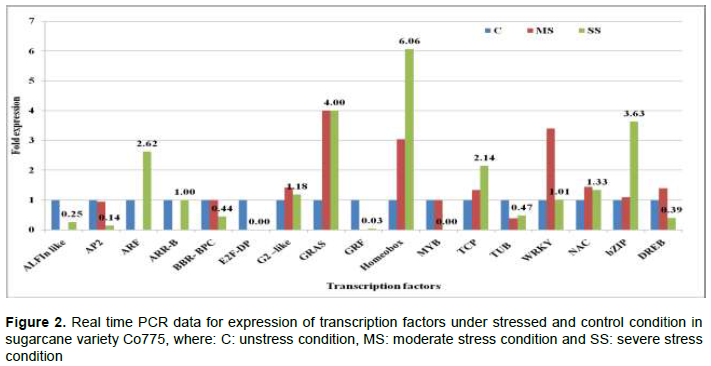
CONCLUSION
CONFLICT OF INTERESTS
ACKNOWLEDGEMENT
REFERENCES
|
Agarwal PK, Jha B (2010). Transcription factors in plants and ABA dependent and independent abiotic stress signaling. Biologia Plantarum 54:201-212. |
|
|
Agustina G, Ferreira TH, Mattos RS, Dias LI, Hoshino AA, Carneiro MS, Souza GM, Tercılio CJr, Nogueira RM, Laurıcio E, Marcelo M (2013). Effects of drought on the microtranscriptome of field-grown sugarcane plants. Planta 237:783-798. |
|
|
Atkin RK, Barton GE, Robinson DK (1973). Effect of root-growing temperature on growth-substances in xylem exudate of Zea mays. Journal of Experimental Botany 24(79):475-487. |
|
|
Bayoumi TY, Eid MH, Metwali EM (2008). Application of physiological and biochemical indices as a screening technique for drought tolerance in wheat genotypes. African Journal of Biotechnology 7:2341-2352. |
|
|
Behnam B, Kikuchi A, Celebi-Toprak F, Yamanaka S, Kasuga M, Yamaguchi-Shinozaki K, Watanabe KN (2006). The Arabidopsis DREB1A gene driven by the stress-inducible rd29A promoter increases salt-stress tolerance in proportion to its copy number intetrasomic tetraploid potato (Solanum tuberosum). Plant Biotechnology 23:169-177. |
|
|
Black CA (1965). Methods of soil analysis: part I physical and mineralogical properties. American Society of Agro, Madison, Wisconsin, USA. |
|
|
Chen H, Lai ZB, Shi JW, Xiao Y, Chen ZX, Xu XP (2010). Roles of Arabidopsis WRKY18, WRKY40 and WRKY60 transcription factors in plant responses to abscisic acid and abiotic stress. BMC Plant Biology 10:281-289. |
|
|
Chen L, Song Y, Li S, Zhang L, Zou C, Yu D (2011). The role of WRKY transcription factors in plant abiotic stresses. Biochimica et Biophysica Acta 36:41-50. |
|
|
Dezar CA, Fedrigo GV, Chan RL (2005). The promoter of the sunflower HD-Zip protein gene HAHB4 directs tissue-specific expression and is inducible by water stress, high salt concentrations and ABA. Plant Sciences 169:447-459. |
|
|
Golldack D, Lüking I, Yang O (2011). Plant tolerance to drought and salinity: stress regulating transcription factors and their functional significance in the cellular transcriptional network. Plant Cell Reports 30:1383-1391. |
|
|
Gomathi R, Vasantha S (2010). Screening for drought tolerance in sugarcane. Extension publication no 180, Sugarcane Breeding institute, Coimbatore, India. |
|
|
Govind G, ThammeGowda HV, Kalaiarasi PJ, Iyer DR, Muthappa SK, Nese S, Makarla UK (2009). Identification and functional validation of a unique set of drought induced genes preferentially expressed in response to gradual water stress in peanut. Molecular Genetics and Genomics 281:591-605. |
|
|
Hemaprabha G, Swapna S (2012). Genetic diversity and selection among drought tolerant genotypes of sugarcane using microsatellite markers. Sugar Tech 14(4):327-333. |
|
|
Hirsch S, Oldroyd GED (2009). GRAS-domain transcription factors that regulate plant development. Plant Signaling and Behavior 4(8):698-700. |
|
|
Hoagland DR, Arnon DI (1950). The water-culture method for growing for plants without soil. Agricultural Experiment Station Circular 347:1- 32. |
|
|
Jain D, Chattopadhyay D (2010). Analysis of gene expression in response to water deficit of chickpea (Cicer arietinum L.) varieties differing in drought tolerance. BMC Plant Biology 10:24. doi:10.1186/1471-2229-10-24. |
|
|
Joshi R, Wani SH, Singh B, Bohra A, Dar ZA, Lone AA, Pareek A, Singla-Pareek SL (2016). Transcription factors and plants response to drought stress: current understanding and future directions. Frontiers in Plant Science 7:1029. |
|
|
Kasuga M, Miura S, Shinozaki K, Yamaguchi-Shinozaki K (2004). A combination of the Arabidopsis DREB1A gene and stress-induciblerd29A promoter improved drought- and low-temperature stress tolerance in tobacco by gene transfer. Plant and Cell Physiology 45:346-350. |
|
|
Kim JH, Choi D, Kende H (2003). The AtGRF family of putative transcription factors is involved in leaf and cotyledon growth in Arabidopsis. The Plant Journal 36(1):94-104. |
|
|
Latchman DS (2003). Eukaryotic transcription factors. Edn 4, San Diego: Academic Press. |
|
|
Ling HQ, Wu G, Jinlong L, Que Y (2014). Comprehensive selection of reference genes for gene expression normalization in sugarcane by real time quantitative RT-PCR. PLOS One 9(5):e97469. |
|
|
Liu Q, Kasuga M, Sakuma Y, Abe H, Miura S, Yamaguchi-Shinozaki K, Shinozaki K (1998). Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively in Arabidopsis. Plant Cell 10:1391-1406. |
|
|
Livak KJ, Schmittgen TD (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCt method. Methods 25:402-408. |
|
|
Lu G, Gao C, Zhong X, Han B (2009). Identification of OsbZIP72 as a positive regulator of ABA response and drought tolerance in rice. Planta 229:605-615. |
|
|
Manel D, Sumangala B (2017). Physiological responses of commercial sugarcane (Saccharum spp. hybrids) varieties to moisture deficit stress tolerance. Indian Journal of Plant Physiology pp. 1-8. |
|
|
Moore PH (1987). Breeding for stress resistance. In: Ed. D.J. Heinz, Sugarcane improvement through breeding, Elsevier, Amsterdam pp. 503-542. |
|
|
Naidu KM (1978). Drought resistance in sugarcane- A review. Biannual conference of Sugarcane Research and Development Workers of Andhra Pradesh. |
|
|
Oh SJ, Song SI, Kim YS, Jang HJ, Kim YS, Kim M, Kim YK, Nahm BH, Kim KJ (2005). Arabidopsis CBF3/DREB1A and ABF3 in transgenic rice increased tolerance to abiotic stress without stunting growth. Plant Physiology 138:341-351. |
|
|
Pasquali G, Biricolti S, Locatelli F, Baldoni E, Mattana M (2008). Osmyb4 expression improves adaptive responses to drought and cold stress in transgenic apples. Plant Cell Reports 27:1677-1686. |
|
|
Qiu YP, Yu DQ (2009). Over-expression of the stress-induced OsWRKY45 enhances disease resistance and drought tolerance in Arabidopsis. Environmental and Experimental Botany 65:35-47. |
|
|
Ramesh P (2000). Effect of different levels of drought during the formative phase on growth parameters and its relationship with dry matter accumulation in sugarcane. Journal of Agronomy and Crop Science 185(2):83-89. |
|
|
Ren XZ, Chen ZZ, Liu Y, Zhang HR, Zhang M, Liu Q, Hong XH, Zhu JK, Gong ZZ (2010). ABO3, a WRKY transcription factor, mediates plant responses to abscisic acid and drought tolerance in Arabidopsis. Plant Journal 63:417-429. |
|
|
Rodrigues FA, Marcelo LL, Zingaretti SM (2009). Analysis of gene expression profiles under water stress in tolerant and sensitive sugarcane plants. Plant Sciences 176:286-302. |
|
|
Sakuma Y, Liu Q, Dubouzet JG, Abe H, Shinozaki K, Yamaguchi-Shinozaki K (2002). DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration-and cold-inducible gene expression. Biochemical and Biophysical Research Communications 290:998-1009. |
|
|
Shang Y, Yan L, Liu ZQ, Zheng C, Chao M, Xin Q, Wu FQ, Wang XF, Du SY, Jiang T, Zhang XF, Zhao R, Sun HL, Liu R, Yu YT, Zhang DP (2010). The Mg-Chelatase H subunit of Arabidopsis antagonizes a group of transcription repressors to relieve ABA-responsive genes of inhibition. Plant Cell 22:1909-1935. |
|
|
Shunwu Y, Fengxian L, Feiming W, Weiwei W, Jiajia L, Hanwei M, Lijun L (2012), Identification of rice transcription factors associated with drought tolerance using the ecotilling method. PLOS One 7(2):e30765. |
|
|
Song Y, Chen LG, Zhang LP, Yu DQ (2010). Over expression of OsWRKY72 gene interferes in the ABA signal and auxin transport pathway of Arabidopsis. Journal of Biosciences 35:459-471. |
|
|
Steppuhn H, Raney JP (2005). Emergence, height, and yield of canola and barley grown in saline root zones. Canadian Journal of Plant Science 85(4):815-827. |
|
|
Swapna S, Hemaprabha G (2010). Identification of two new drought specific candidate genes in sugarcane (Saccharum spp.). Electronic Journal of Plant Breeding 1(4):1164-1170. |
|
|
Tran LS, Nakashima K, Sakuma Y, Simpson SD, Fujita Y (2004). Isolation and functional analysis of Arabidopsis stress-inducible NAC transcription factors that bind to a drought-responsive cis-element in the early responsive to dehydration stress 1 promoter. Plant Cell 16:2481-2498. |
|
|
Uno Y, Furihata T, Abe H, Yoshida R, Shinozaki K, Yamaguchi-Shinozaki K (2000). Arabidopsis basic leucine zipper transcription factors involved in an abscisic acid-dependent signal transduction pathway under drought and high-salinity conditions. Proceedings of the National Academy of Sciences of the United States of America 97:11632-11637 |
|
|
Vannini C, Locatelli F, Bracale M, Magnani E, Marsoni M, Osnato M, Mattana M, Baldoni E, Coraggio S (2004). Over expression of the rice Osmyb4 gene increases chilling and freezing tolerance of Arabidopsis thaliana plants. Plant Journal 37:115-127. |
|
|
Vannini M, Campa M, Iriti M, Genga A, Faoro F, Carravieri S, Rotino SL, Rossoni M, Spinardi A, Bracale M (2007). Evaluation of transgenic tomato plants ectopically expressing the rice Osmyb4 gene. Plant Sciences 173:231-239. |
|
|
Waclawovsky AJ, Sato PM, Lembke CG, Moore PH, Souza GM (2010). Sugarcane for bioenergy production:an assessment of yield and regulation of sucrose content. Plant Biotechnology Journal 8(3):263-276. |
|
|
Wu X, Shiroto Y, Kishitani S, Ito Y, Toriyama K (2009). Enhanced heat and drought tolerance in transgenic rice seedlings overexpressing OsWRKY11 under the control of HSP101 promoter. Plant Cell Reports 28:21-30. |
|
|
Xiong L, Zhu JK (2002). Molecular and genetic aspects of plant responses to osmotic stress. Plant, Cell and Environment 25:131-139. |
|
|
Xu K, Chen S, Li T, Ma X, Liang X, Ding X, Liu H, Luo L (2015). OsGRAS23, a rice GRAS transcription factor gene, is involved in drought stress response through regulating expression of stress-responsive genes. BMC Plant Biology 15:141. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


