Full Length Research Paper

ABSTRACT
Human interleukin 2 protein (IL-2) is an important cytokine found to be elevated in several types of cancer. A synthetic DNA sequence of the cDNA of mature IL-2 protein was cloned into the pRSET-B expression vector. The expressed IL-2 protein in Escherichia coli [BL21] was associated with the formation of insoluble inclusion bodies (IBs). The effect of different cultivation conditions (temperature, isopropyl-β-D-thiogalactoside concentration, and early harvest of cells) together with the incorporation of single or dual His-tag on the formation of IBs of the expressed protein was investigated. Yet, expression of soluble IL-2 was not achieved under any of the investigated conditions. A simple protocol for rapid and effective solubilization of these IBs was optimized. Using this protocol, together with subsequent purification using ion metal affinity chromatography, a purified His-IL2 protein was obtained in a yield of 5.1 mg/cell pellet of 1 L culture. In conclusion, the effect of different expression conditions on the solubility behavior of an expressed eukaryotic protein in E. coli was investigated using human IL-2 as a model protein. Moreover, the purified expressed protein could be used as a positive control in early diagnosis of tumors and in cancer research in Egypt.
Key words: Human interleukin-2, protein expression, inclusion bodies.
INTRODUCTION
MATERIALS AND METHODS

RESULTS
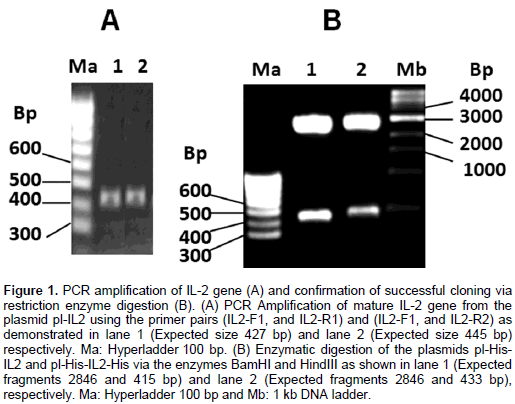
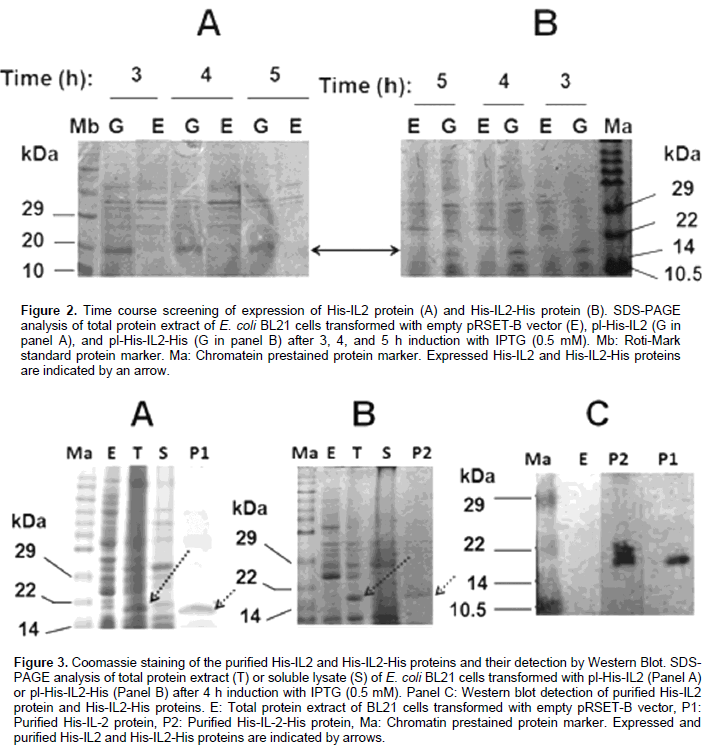
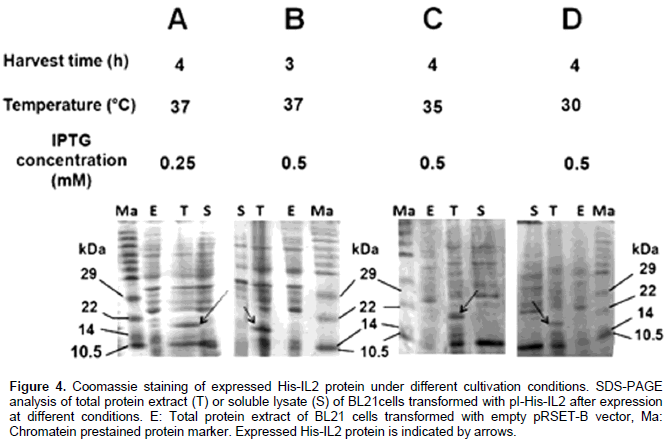
DISCUSSION
CONCLUSION
CONFLICT OF INTERESTS
REFERENCES
|
Akdis M, Aab A, Altunbulakli C, Azkur K, Costa RA, Crameri R, Duan S, Eiwegger T, Eljaszewicz A, Ferstl R, Frei R (2016). Interleukins (from IL-1 to IL-38), interferons, transforming growth factor beta, and TNF-alpha: Receptors, functions, and roles in diseases. J. Allergy Clin. Immunol. 138(4):984-1010. |
|
|
Cloeckaert A, Baucheron S, Vizcaino N, Zygmunt MS (2001). Use of recombinant BP26 protein in serological diagnosis of Brucella melitensis infection in sheep. Clin. Diagn. Lab. Immunol. 8(4):772-775. |
|
|
Costa S, Almeida A, Castro A, Domingues L (2014). Fusion tags for protein solubility, purification and immunogenicity in Escherichia coli: the novel Fh8 system. Front. Microbiol. 5:63. |
|
|
Devos R, Plaetinck G, Cheroutre H, Simons G, Degrave W, Tavernier J, Remaut E, Fiers W (1983). Molecular cloning of human interleukin 2 cDNA and its expression in E. coli. Nucleic Acids Res. 11(13):4307-4323. |
|
|
El-Mowafy M, Bahgat MM, Bilitewski U (2013). Deletion of the HAMP domains from the histidine kinase CaNik1p of Candida albicans or treatment with fungicides activates the MAP kinase Hog1p in S. cerevisiae transformants. BMC Microbiol. 13:209. |
|
|
Garcia-Tunon I, Ricote M, Ruiz A, Fraile B, Paniagua R, Royuela M (2004). Interleukin-2 and its receptor complex (alpha, beta and gamma chains) in in situ and infiltrative human breast cancer: an immunohistochemical comparative study. Breast Cancer Res. 6(1):R1-7. |
|
|
Gatti-Lafranconi P, Natalello A, Ami D, Doglia SM, Lotti M (2011). Concepts and tools to exploit the potential of bacterial inclusion bodies in protein science and biotechnology. FEBS J. 278(14):2408-2418. |
|
|
Gehman LO, Robb RJ (1984). An ELISA-based assay for quantitation of human interleukin 2. J. Immunol. Methods 74(1):39-47. |
|
|
Janik JE, Sharfman WH, Smitht JW, Longo DL, Sznol M, Urba WJ, Figlin R, Pierce WC, Bukowski RM, Fyfe G, Belldegrun A (1993). Erythropoietin production. A potential marker for interleukin-2/interferon-responsive tumors. Cancer 72(9):2656-2659. |
|
|
Ju G, Collins L, Kaffka KL, Tsien WH, Chizzonite R, Crowl R, Bhatt R, Kilian PL (1987). Structure-function analysis of human interleukin-2. Identification of amino acid residues required for biological activity. J. Biol. Chem. 262(12):5723-5731. |
|
|
Laemmli UK (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227(5259):680-685. |
|
|
Lee MS, Hseu YC, Lai GH, Chang WT, Chen HJ, Huang CH, Lee MS, Wang MY, Kao JY, You BJ, Lin WH (2011). High yield expression in a recombinant E. coli of a codon optimized chicken anemia virus capsid protein VP1 useful for vaccine development. Microb. Cell Fact. 10:56. |
|
|
Lin WC, Yasumura S, Suminami Y, Sung MW, Nagashima S, Stanson J, Whiteside TL (1995). Constitutive production of IL-2 by human carcinoma cells, expression of IL-2 receptor, and tumor cell growth. J. Immunol. 155(10):4805-4816. |
|
|
Ma X, Zheng W, Wei D, Ma Y, Wang T, Wang J, Liu Q, Yang S (2006). Construction, expression, and purification of HIV-TAT-survivin (T34A) mutant: a pro-apoptosis protein in Escherichia coli. Protein Expr. Purif. 47(1):36-44. |
|
|
Palmer I, Wingfield PT (2004). Preparation and extraction of insoluble (inclusion-body) proteins from Escherichia coli. Current protocols in protein science, Chapter 6: Unit 63. |
|
|
Reichert TE, Watkins S, Stanson J, Johnson JT, Whiteside TL (1998). Endogenous IL-2 in cancer cells: a marker of cellular proliferation. J. Histochem. Cytochem. 46(5):603-611. |
|
|
Ridings J, Macardle PJ, Byard RW, Skinner J, Zola H (1995). Cytokine receptor expression by solid tumours. Ther. Immunol. 2(2):67-76. |
|
|
Robb RJ, Kutny RM, Chowdhry V (1983). Purification and partial sequence analysis of human T-cell growth factor. Proc. Natl. Acad. Sci. USA, 80(19):5990-5994. |
|
|
Roediger B, Kyle R, Tay SS, Mitchell AJ, Bolton HA, Guy TV, Tan SY, Forbes-Blom E, Tong PL, Köller Y, Shklovskaya E (2015). IL-2 is a critical regulator of group 2 innate lymphoid cell function during pulmonary inflammation. J. Allergy Clin. Immunol. 136(6):1653-1663. |
|
|
Rosano GL, Ceccarelli EA (2014). Recombinant protein expression in Escherichia coli: advances and challenges. Front. Microbiol. 5:172. |
|
|
Rosenberg SA, Grimm EA, Mcgrogan M, Doyle M, Kawasaki E, Koths K, Mark DF (1984). Biological activity of recombinant human interleukin-2 produced in Escherichia coli. Science 223(4643):1412-1414. |
|
|
Royuela M, De Miguel MP, Bethencourt FR, Fraile B, Arenas MI, Paniagua R (2000). IL-2, its receptors, and bcl-2 and bax genes in normal, hyperplastic and carcinomatous human prostates: immunohistochemical comparative analysis. Growth Factors 18(2):135-146. |
|
|
Schoner RG, Ellis LF, Schoner BE (1992). Isolation and purification of protein granules from Escherichia coli cells overproducing bovine growth hormone. 1985. Biotechnology (Reading, Mass.) 24:349-352. |
|
|
Sengupta P, Meena K, Mukherjee R, Jain SK, Maithal K (2008). Optimized conditions for high-level expression and purification of recombinant human interleukin-2 in E. coli. Indian J. Biochem. Biophys. 45(2):91-97. |
|
|
Shaaban MI, Abdelmegeed E, Ali YM (2015). Cloning, Expression, and Purification of Recombinant Uricase Enzyme from Pseudomonas aeruginosa Ps43 Using Escherichia coli. J. Microbiol. Biotechnol. 25(6):887-892. |
|
|
Singh A, Upadhyay V, Upadhyay AK, Singh SM, Panda AK (2015). Protein recovery from inclusion bodies of Escherichia coli using mild solubilization process. Microb. Cell Fact. 14:41. |
|
|
Smith GE, Ju G, Ericson BL, Moschera J, Lahm HW, Chizzonite R, Summers MD (1985). Modification and secretion of human interleukin 2 produced in insect cells by a baculovirus expression vector. Proc. Natl. Acad. Sci. USA. 82(24):8404-8408. |
|
|
Sorensen HP, Mortensen KK (2005a). Soluble expression of recombinant proteins in the cytoplasm of Escherichia coli. Microb. Cell Fact. 4(1):1. |
|
|
Sorensen HP, Mortensen KK (2005b). Advanced genetic strategies for recombinant protein expression in Escherichia coli. J. Biotechnol. 115(2):113-128. |
|
|
Stern AS, Pan YC, Urdal DL, Mochizuki DY, Dechiara S, Blacher R, Wideman J, Gillis S (1984). Purification to homogeneity and partial characterization of interleukin 2 from a human T-cell leukemia. Proc. Natl. Acad. Sci. USA. 81(3):871-875. |
|
|
Taniguchi T, Matsui H, Fujita T, Takaoka C, Kashima N, Yoshimoto R, Hamuro J (1983). Structure and expression of a cloned cDNA for human interleukin-2. Nature 302(5906):305-310. |
|
|
Uma M, Rao PP, Nagalekshmi K, Hegde NR (2016). Expression and purification of polioviral proteins in E. coli, and production of antisera as reagents for immunological assays. Protein Expr. Purif. 128:115-122. |
|
|
Wang X, Liu X, Zhang Y, Wang Z, Zhu G, Han G, Chen G, Hou C, Wang T, Ma N, Shen B (2016). Interleukin (IL)-39 [IL-23p19/Epstein-Barr virus-induced 3 (Ebi3)] induces differentiation/expansion of neutrophils in lupus-prone mice. Clin. Exp. Immunol. 186(2):144-156. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


