Review

ABSTRACT
The importance of biotechnology involved in the availability of nutrients to plants in different production systems is well known. In the search for agricultural sustainability, biological nitrogen fixation process stands out, especially in tropical regions where soil organic matter can be rapidly mineralized. In this aspect, researches have pointed out the potentialities of the use of diazotrophic bacteria, as well as other growth-promoting bacteria in Poaceae. Maize crop, especially, stands out in the international scenario, requiring a deepening of the research aiming to raise the contribution potential of microorganisms including Azospirillum spp. in reducing the consumption of fertilizers from non-renewable sources while promoting an increase in agricultural productivity and mitigating environmental impacts.
Key words: Zea mays, diazotrophic bacteria, nitrogen, biological fixation, growth promoting.
INTRODUCTION
MAIZE CROP
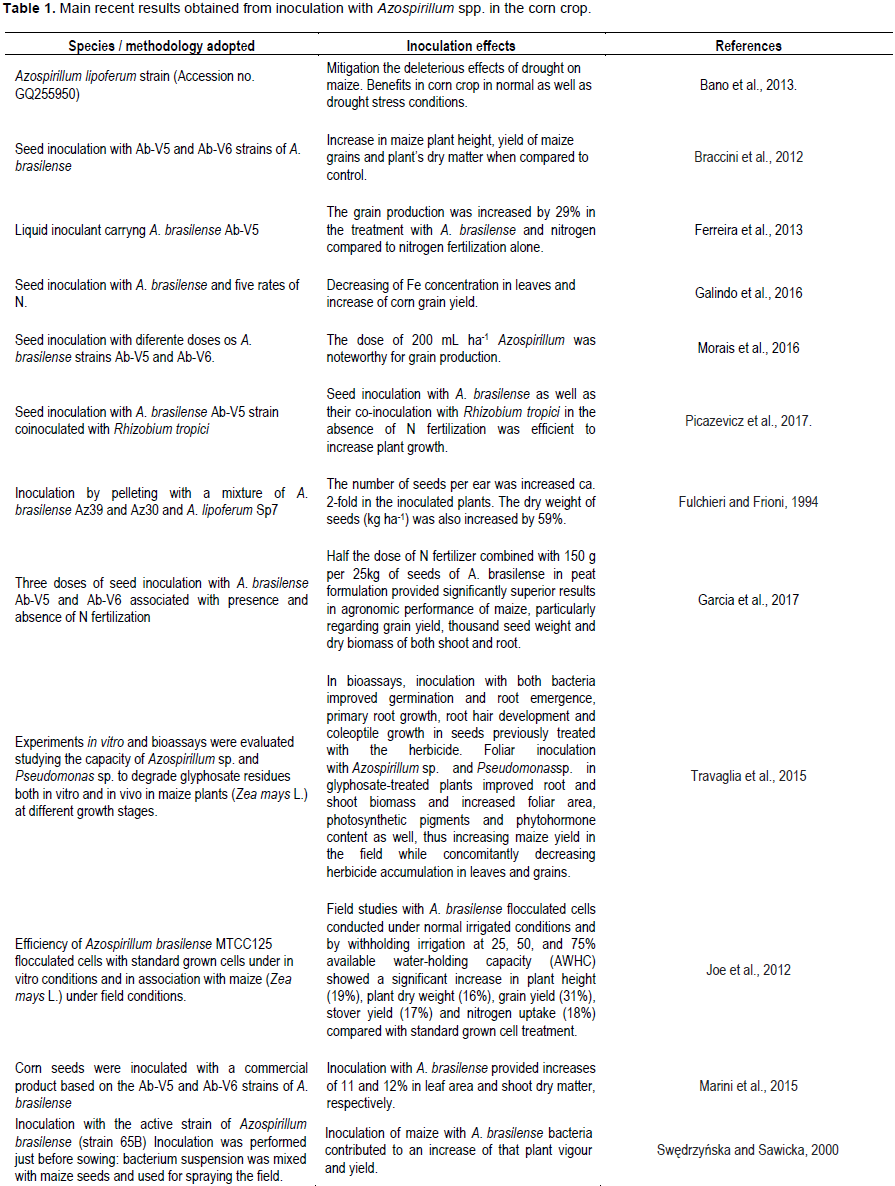

FINAL CONSIDERATIONS
CONFLICT OF INTERESTS
REFERENCES
|
ANDA - Associação Nacional para Difusão de Adubos (2017). Mercado de Fertilizantes. Jan-Sep 2017. |
|
|
Araújo RM, Araújo ASF, Nunes LAPL, Figueiredo MVB (2014). Resposta do milho verde à inoculação com Azospirillum brasilense e níveis de nitrogênio. Cienc. Rural 44(9):1556-1560. |
|
|
Ardakani MR, Mazaheri D, Mafakheri S, Moghaddam A (2011). Absorption efficiency of N, P, K through triple inoculation of wheat (Triticum aestivum L.) by Azospirillum brasilense, Streptomyces sp., Glomus intraradices and manure application. Physiol. Mol. Biol. Plants 17:181-192. |
|
|
Argenta G, Silva PRF, Sangoi L (2001). Arranjo de plantas em milho: análise do estado-da-arte. Cienc. Rural 31(6):1075-1084. |
|
|
Baldani JI, Caruso L, Baldani VLD, Goi SR, Döbereiner J (1997). Recent advances in BFN with non-legume plants. Soil Biol. Biochem. 29:911-922. |
|
|
Bano Q, Ilyas N, Bano A, Zafar N, Akram A, Hassan F (2013). Effect of Azospirillum inoculation on maize (Zea mays L.) under drought stress. Pak. J. Bot. 45:13-20. |
|
|
Barassi CA, Sueldo RJ, Creus CM, Carrozzi LE, Casanovas WM, Pereyra MA (2008). Potencialidad de Azospirillum en optimizer el crecimiento vegetal bajo condiciones adversas. In: Cassán FD, García de Salamone I. Azospirillum spp.: cell physiology, plant interactions and agronomic research in Argentina. Asociación Argentina de Microbiologia, pp. 49-59. |
|
|
Bárbaro IM, Brancalião SR, Ticelli M (2008). É possível fixação biológica de nitrogênio no milho. INFOBIBOS - Tecnological informations. Online. |
|
|
Barros JFC, Calado JG (2014). A cultura do milho. Évora: Univerisade de Évora. 52 p. |
|
|
Barros Neto CR (2008). Efeito do nitrogênio e da inoculação de sementes com Azospirillum brasilense no rendimento de grãos de milho. Ponta Grossa: Universidade Estadual de Ponta Grossa. 29 p. |
|
|
Bartchechen A, Fiori CCL, Watanabe SH, Guarido RC (2010). Efeito da inoculação de Azospirillum brasilense na produtividade da cultura do milho (Zea mays L.). Campo Digit. 5(1):56-59. |
|
|
Bashan Y, Bashan LE (2005). Pant growth-promoting. In. Hillel D (ed) Encyclopedia of soils in the environment, Elsevier, Oxford, UK, 1:103-115. |
|
|
Bashan Y, De-Bashan LE (2010). How the plant growth-promoting bacterium Azospirillum promotes plant growth - a critical assessment. Adv. Agron. 108:77-136. |
|
|
Bashan Y, Holguin G, De-Bashan LE (2004). Azospirillum-plant relations physiological, molecular, agricultural, and environmental advances. Can. J. Microbiol. 50:521-577. |
|
|
Bashan Y, Levanony H (1990). Current status of Azospirillum inoculation technology: Azospirillum as a challenge for agriculture. Can. J. Microbiol. 36:591-608. |
|
|
Bashan Y, Puente ME, Rodriguez-Medonza MN, Toledo G, Holguin G, Ferrera-Cerrato R, Pedrin S (1995). Survival of Azospirillum brasilense in the bulk soil and rhizosphere of 23 soil types. Appl. Environ. Microbiol. 61:1938-1945. |
|
|
Basi S (2013). Associação de Azospirillum brasilense e de nitrogênio em cobertura na cultura do milho. Guarapuava: Universidade Estadual do Centro-Oeste. 50 p. |
|
|
Basi S, Lopes ECP, Kaminski TH, Pivatto RAD, Cheng NC, Sandini IE (2011). Azospirillum brasilense nas sementes e no sulco de semeadura da cultura do milho. In. Semana de Integração, Ensino, Pesquisa e Extensão. Guarapuava: II SIEPE. |
|
|
Bottini R, Fulchieri M, Pearce D, Pharis R (1989). Identification of gibberellins A1, A3, and iso-A3 in cultures of A. lipoferum. Plant Physiol. 90:45-47. |
|
|
Braccini AL, Dan LGM, Piccinin GG, Albrecht LP, Barbosa MC, Ortiz AHT (2012). Seed inoculation with Azospirillum brasilense, associated with the use of bio-regulators in maize. Rev. Caatinga 25(2):58-64. |
|
|
Cáceres EAR (1982). Improved medium for isolation of Azospirillim spp. Appl. Environ. Microbiol. 44(43):990-991. |
|
|
Campos BHC, Thiesen S, Gnatta V (1999). Inoculante "Graminante" nas culturas de trigo e aveia. Cienc. Rural 23(3):401-407. |
|
|
Cantarella H (1993). Calagem e adubação do milho. In. Büll LT, Cantarella H (eds) Cultura do milho: fatores que afetam a produtividade. Piracicaba: Potafos. pp. 147-185. |
|
|
Cassán F, Sgroy V, Perrig D, Masciarelli O, Luna V (2008). Producción de fitohormonas por Azospirillum sp. Aspectos fisiológicos y tecnológicos de la promoción del crecimiento vegetal. In: Cassán FD, García de Salamone I (eds) Azospirillum spp.: cell physiology, plant interactions and agronomic research in Argentina. Argentina: Asociación Argentina de Microbiologia. pp. 61-86. |
|
|
Cassán FD, García de Salamone I (2008). (eds) Azospirillum spp.: cell physiology, plant interactions and agronomic research in Argentina. Argentina: Asociación Argentina de Microbiologia. 268 p. |
|
|
Cavallet LE, Pessoa ACS, Helmich JJ, Helmich PR, Ost CF (2000) Produtividade do milho em resposta à aplicação de nitrogênio e inoculação das sementes com Azospirillum spp. Rev. Bras. Eng. Agríc. Ambient. 4:129-132. |
|
|
Chavarria G, De Mello N (2011). Bactérias do gênero Azospirillum e sua relação com gramíneas. Rev. Plantio Direto, 125. |
|
|
Cheng NC, Novakowiski JH, Sandini I, Domingues L (2011). Substituição da adubação nitrogenada de base pela inoculação com Azospirillum brasilense na cultura do milho. In. Seminário Nacional de Milho Safrinha. Lucas do Rio Verde: Fundação Rio Verde. pp. 377-382. |
|
|
Companhia Nacional de Abastecimento (CONAB) (2017). Acompanhamento da safra brasileira de grãos. 2017/2018 Season. |
|
|
Correa OS, Romero AM, Soria MA, De Estrada M (2008). Azospirillum brasilense-plant genotype interactions modify tomato response to bacterial diseases, and root and foliar microbial communities. In: Cassán FD, Garcia De Salamone I (eds) Azospirillum spp.: cell physiology, plant interactions and agronomic research in Argentina. Argentina: Associación Argentina de Microbiologia. pp. 87-95. |
|
|
Costa RRG, Quirino GSF, Naves DCF, Santos CB, Rocha AFS (2015). Efficiency of inoculant with Azospirillum brasilense on the growth and yield of second-harvest maize. Pesq. Agropecu. Trop. 45(3):304-311. |
|
|
Croes CL, Moens S, Van Bastelaere E, Vanderleyden J, Michiels KW (1993). The polar flagellum mediates Azospirillum brasilense adsorption to wheat roots. J. Gen Microbiol. 139:2261-2269. |
|
|
Cruz JC, Pereira Filho IA, Garcia JC, Duarte JO (2010). Cultivo do Milho: Cultivares. Sistema de Produção. 6ed. Sete Lagoas: Embrapa Milho e Sorgo. 8 p. |
|
|
Cunha FN, Silva NF, Bastos FJC, Carvalho JJ, Moura LMF, Teixeira MB, Rocha AC, Souchie EL (2014). Efeito da Azospirillum brasilense na Produtividade de Milho no Sudoeste Goiano. Rev. Bras. Milho Sorgo. 13:261-272. |
|
|
De Salamone IG, Dobereiner J, Urquiaga S, Boddey RM (1996). Biological nitrogen fixation in Azospirillum strain-maize genotype associations as evaluated by the 15 N isotope dilution technique. Biol Fertil. Soils 23(3):249-256. |
|
|
Del Gallo M, Fendrik I (1994). The rhizosphere and Azospirillum. In. Okon Y (eds) Azospirillum plant association. Boca Raton: CGC Critical Reviews in Plant Science. pp. 57-75. |
|
|
Díaz-Zorita M, Fernandez Canigia MV (2008). Análisis de la producción de cereales inoculados con Azospirillum brasilense en la República Argentina. In. Cassán FD, Garcia de Salamone I (eds) Azospirillum spp.: cell physiology, plant interactions and agronomic research in Argentina. Argentina: Asociación Argentina de Microbiologia. pp. 155-166. |
|
|
Döbereiner J, Baldani VLD, Baldani JI (1995). Como isolar e identificar bactérias diazotróficas de plantas não-leguminosas. Brasília: Embrapa-SPI. 60 p. |
|
|
Döbereiner J, Pedrosa FO (1987). Nitrogen-fixing bacteria in non-leguminous crop plants. Science Tech, Madison, USA. 155 p. |
|
|
Döbereiner, J (1992). History and new perspectives of diazotrophs in association with non-leguminous plants. Symbiosis. 13:1-13. |
|
|
Duarte AP, Piedade RC, Martins VC, Cantarella H, Barros VLNP (2012). Resposta de cultivares de milho ao nitrogênio em cobertura e à inoculação com Azospirillum. In. 29 Congresso nacional de milho e sorgo. Campinas: Instituto Agronômico/Associação Brasileira de Milho e Sorgo. pp. 1786-1792. |
|
|
Epstein E, Bloom AJ (2006). Nutrição mineral de plantas: princípios e perspectivas. Londrina: Editora Planta. 403 p. |
|
|
Fancelli AL (1997). Cultura do milho: A importância da tecnologia. Informações Agronômicas, Piracicaba. 1(78):4-6. |
|
|
FAO– Food and Agriculture Organization Statistical Databases (2017). Crops. FAO-Food and Agriculture Organization of the United Nations. |
|
|
Farinelli R, Hanashiro RK, Amaral CB, Fornasieri Filho D (2012). Reposta da cultura do milho à inoculação de sementes e adubação nitrogenada em cobertura. In. 29 Congresso nacional de milho e sorgo. Campinas: Instituto Agronômico/Associação Brasileira de Milho e Sorgo. pp. 1672-1678. |
|
|
Ferreira AS, Pires RR, Rabelo PG, Oliveira RC, Luz JMQ, Brito CH (2013). Implications of Azospirillum brasilense inoculation and nutrient addition on maize in soils of the Brazilian Cerrado under greenhouse and field conditions. Appl. Soil Ecol. 72:103-108. |
|
|
Fornasieri Filho D (2007). Manual da cultura do milho. Jaboticabal: Funep. 574 p. |
|
|
Fukami J, Nogueira MA, Araujo RS, Hungria M (2016). Acessing inoculation methods of maize and wheat with Azospirillum brasilense. AMB Express 6:1-13. |
|
|
Fulchieri M, Frioni L (1994). Azospirillum inoculation on maize (Zea mays): effect on yield in a field experiment in central Argentina. Soil Biol. Biochem. 26(7):921-923. |
|
|
Galindo FS, Teixeira Filho MCM, Buzetti S, Santini JMK, Alves CJ, Nogueira LM, Ludkiewicz MGZ, Andreotti M, Bellotte JLM (2016). Corn yield and foliar diagnosis affected by nitrogen fertilization and inoculation with Azospirillum brasilense. Rev. Bras. Ciênc. Solo 40:1-18. |
|
|
García de Salomone, Döbereiner J (1996). Maize genotype effects on the response to Azospirillum inoculation. Biol. Fertil. Soils 21(3):193-196. |
|
|
Garcia MM, Pereira LC, Braccini AL, Angelotti P, Suzukawa AK, Marteli DC, Felber PH, Bianchessi PA, Dametto IB (2017). Effects of Azospirillum brasilense on growth and yield compounds of maize grown at nitrogen limiting conditions. Rev. Ciênc Agrár. 40(2):353-362. |
|
|
Grassi Filho H (2010). Funções do nitrogênio e enxofre nas plantas. In. Vale DW, Sousa JI, Prado RM (eds) Manejo da fertilidade do solo e nutrição de plantas. Jaboticabal: FCAV, pp. 187-198. |
|
|
Hall PG, Krieg NR (1984). Application of the indirect immunoperoxidase stain technique to the flagella of Azospirillum brasilense. Appl. Environ. Microbiol. 47(2):433-435. |
|
|
Hungria M (2011). Inoculação com Azospirillum brasilense: inovaçãoem rendimento a baixo custo. Londrina: Embrapa. |
|
|
Hungria M, Campo RJ, Souza EMS, Pedrosa FO (2010). Inoculation with selected strains of Azospirillum brasilense and A. lipoferum improves yields of maize and wheat in Brazil. Plant Soil 331(1/2):413-425. |
|
|
Joe MM, Karthikeyan B, Chauhan PS, Shagol C, Islam MR, Deiveekasundaram M, Sa T (2012). Survival of Azospirillum brasilense flocculated cells in alginate and its inoculation effect on growth and yield of maize under water deficit conditions. Eur. J. Soil Biol. 50:198-206. |
|
|
Kannan T, Ponmurugan P (2010). Response of paddy (Oryza sativa L.) varieties to Azospirillum brasilense inoculation. J. Phytol. 2:8-13. |
|
|
Kennedy IR, Choudhury ATMA, Kecskes ML (2004). Non-symbiotic bacterial diazotrophs in crop-farming systems: can their potential for plant growth promotion be better explored. Soil Biol. Biochem. 36:1229-1244. |
|
|
Lange A, Moreira FMA (2002). Detecção de Azospirillum amazonense em raízes e rizosfera de Orchidaceae e de outras famílias vegetais. Rev. Bras. Ciênc. Solo 26:535-543. |
|
|
Lewis WM, Saunders JF, Crumpaker DW, Brendecke CM (1984). Eutrophication and land use, Lake Dillon, Colorado. Ecol. Stud. 46:202. |
|
|
Lindemann WC, Glover CR (2003). Nitrogen fixation by Legumes. Cooperative extension service. New México State University: College of Agriculture and Homes Economics. Guide A-129. |
|
|
Magalhães FMM, Döbereiner J (1984). Ocorrência de Azospirillum amazonense em alguns ecossistemas da Amazônia. Rev. Microbiol. 15:246-252. |
|
|
Malavolta E (2006). Manual de nutrição mineral de plantas. Piracicaba: Ceres. 631 p. |
|
|
Marini D, Guimarães VF, Dartora J, Lana MC, Pinto Júnior AS (2015). Growth and yield of corn hybrids in response to association with Azospirillum brasilense and nitrogen fertilization. Rev. Ceres. 62(1):117-123. |
|
|
Marschner H (1995). Mineral nutrition of higher plants. London: Academic Press. 889 p. |
|
|
Martínez-Morales LJ, Soto-Urzúa L, Baca BE, Sánchez-Ahédo JA (2003). Indole-3-butyric acid (IBA) production in culture medium by wild strain Azospirillum brasilense. FEMS Microbiol. Lett. 228:167-173. |
|
|
Martins FAD, Andrade AT, Condé ABT, Godinho DB, Caixeta CG, Costa RL, Pomela AWV, Soares CMS (2012). Avaliação de híbridos de milho inoculados com Azospirillum brasilense. Pesq. Agropecu. Gau. 18(2):113-128. |
|
|
Morais TP, Brito CH, Brandão AM, Rezende WS (2016). Inoculation of maize with Azospirillum brasilense in the seed furrow. Rev. Cienc. Agron. 47(2):290-298. |
|
|
Moreira FMS, Da Silva K, Nobrega RSA, Carvalho F (2010). Bactérias diazotróficas associativas: diversidade, ecologia e potencial de aplicações. Comunicata Scientiae 1(2):74-99. |
|
|
Moreira FMS, Siqueira JO (2006). Microbiologia e bioquímica do solo. 2ed. Lavras: UFLA. 729 p. |
|
|
Môro GV, Fritsche-Neto R (2015). Importância e usos do milho no Brasil. In. Borém A, Galvão JCC, Pimentel MA (eds) Milho do plantio à colheita. Viçosa: UFV. pp. 9-23. |
|
|
Müller TM, Bazzanezi AN, Vidal V, Turok JDN, Rodrigues JD, Sandini IE (2012). Inoculação de Azospirillum brasilense no tratamento de sementes e sulco de semeadura na cultura do milho. In. 29 Congresso nacional de milho e sorgo. Campinas: Instituto Agronômico/Associação Brasileira de Milho e Sorgo. pp. 1665-1671. |
|
|
Müller TM, Sandini IE, Rodrigues JD, Novakowiski JH, Basi S, Kaminski TH (2016). Combination of inoculation methods of Azospirilum brasilense with broadcasting of nitrogen fertilizer increases corn yield. Cienc. Rural 46(2):210-215. |
|
|
Nehl DB, Allem SJ, Brown JF (1996). Deleterious rhizosphere bacteria: an integrating perspective. Appl. Soil Ecol. 5:1-20. |
|
|
Novakowiski JH, Sandini IE, Falbo MK, Moraes A, Novakowiski JH, Cheng NC (2011). Efeito residual da adubação nitrogenada e inoculação de Azospirillum brasilense na cultura do milho. Semina: Ciênc. Agrár. 32(1):1687-1698. |
|
|
Okon Y, Labandera-Gonzales CA (1994). Agronomic applications of Azospirillum: an evaluation of 20 years worldwide field inoculation. Soil Biol Biochem. 26(12):1591-1601. |
|
|
Okon Y, Vanderleyden J (1997). Root-associated Azospirillum species can stimulate plants. Appl. Environ. Microbiol. 6(7):366-370. |
|
|
Paterniani E, Campos MS (1999). Melhoramento do milho. In. Borém A (eds) Melhoramento de espécies cultivadas. Viçosa: UFV. p. 817. |
|
|
Pedrinho EAN (2009). Isolamento e caracterização de bactérias promotoras de crescimento em milho (Zea mays L.). Jaboticabal: Faculdade de Ciências Agrárias e Veterinárias, Universidade Estadual Paulista "Júlio de Mesquita Filho". p. 87. |
|
|
Pereira LM, Pereira EM, Revolti LTM, Zingaretti SM, Moro GV (2015). Seed quality, chlorophyll content index and leaf nitrogen levels in maize inoculated with Azospirillum brasilense. Rev. Cienc. Agron. 46:630-637. |
|
|
Perrig D, Boiero L, Masciarelli O, Penna C, Cassán F, Luna V (2007). Plant growth promoting compounds produced by two agronomically important strains of Azospirillum brasilense, and their implications for inoculant formulation. Appl. Microbiol. Biotechnol. 75:1143-1150. |
|
|
Picazevicz AA, Kusdra JF, Moreno ADL (2017). Maize growth in response to Azospirillum brasilense, Rhizobium tropici, molybdenum and nitrogen. Rev. Bras. Eng. Agríc. Ambient. 21(9):623-627. |
|
|
Portugal JR, Arf O, Peres AR, De Castilho Gitti D, Garcia NFS (2017). Coberturas vegetais, doses de nitrogênio e inoculação com Azospirillum brasilense em milho no Cerrado. Rev. Cienc. Agron. 48(4):639-649. |
|
|
Portugal JR, Arf O, Peres AR, Gitti DC, Rodrigues RAF, Garcia NFS, Gare LM (2016). Azospirillum brasilense promotes increment in corn production. Afr. J. Agric. Res. 11:1688-1698. |
|
|
Quadros PD (2009). Inoculação de Azospirillum spp. em sementes de genótipos de milho cultivados no Rio Grande do Sul. Porto Alegre: Universidade Federal do Rio Grande do Sul. p. 74. |
|
|
Reis Júnior FB, Mendes IC, Reis VM, Hungria, M (2010). Fixação biológica de nitrogênio: uma revolução na agricultura. In. Faleiro FG, Andrade SRM, Reis Junior, FB (eds) Estado da arte e aplicações na agropecuária. Planaltina: Embrapa. pp. 247-281. |
|
|
Reis Júnior FB, Reis VM, Silva LG, Döbereiner J (2000). Levantamento e quantificação de bactérias diazotróficas em diferentes genótipos de cana-de-açúcar (Saccharum spp.). Pesq. Agrop. Bras. 35:985-994. |
|
|
Revolti LTM (2014). Interação genótipo vs formas de inoculação com Azospirillum brasilense em milho. Jaboticabal: Universidade Estadual Paulista "Júlio de Mesquita Filho", Faculdade de Ciências Agrárias e Veterinárias. 46 p. |
|
|
Ritchie SW, Hanway JJ, Benson GO (1993). How a Corn Plant Develops, Special Report, n.48, Iowa State University of Science and Technology, Ames, Iowa. |
|
|
Rodriguez H, Gonzalez T, Goire I, Bashan Y (2004). Gluconic acid production and phosphate solubilization by the plant growth-promoting bacterium Azospirillum spp. Naturwissenschaften 91:552-555. |
|
|
Rudnik P, Meletzus D, Green A, He L, Kennedy C (1997). Regulation of nitrogen fixation by ammonium in diazotrophic species of proteobacteria. Soil Biol. Biochem. 29:831-841. |
|
|
Saikia SP, Bora D, Goswami A, Mudoi KD, Gogoi AA (2012). Review on the role of Azospirillum in the yield improvement of non-leguminous crops. Afr. J. Microbiol. Res. 6(6):1085-1102. |
|
|
Sala VMR, Cardoso EJBN, Freitas JG, Silveira APD (2008). Novas bactérias diazotróficas endofíticas na cultura do trigo em interação com a adubação nitrogenada, no campo. Rev. Bras. Cienc. Solo. 32(3):1099-1106. |
|
|
Silva AP, Arruda TF, Bach EE (2004). Ação do Azopirillum brasilense no desenvolvimento das plantas de trigo (variedade IAC-24) e cevada (variedade CEV 95033). Conscientiae Saúde 3:29-35. |
|
|
Strzelczyk E, Kamper M, Li C (1994). Cytokinin-like-substances and ethylene production by Azospirillum in media with different carbon sources. Microbiol. Res. 149:55-60. |
|
|
Swędrzyńska D, Sawicka A (2000). Effect of inoculation with Azospirillum brasilense on development and yielding of maize (Zea mays ssp. saccharata L.) under different cultivation conditions. Pol. J. Environ. Stud. 9(6):505-509. |
|
|
Taiz L, Zeiger E (2013). Fisiologia vegetal. 5ed. Porto Alegre: Artmed,. 954 p. |
|
|
Tien TM, Gaskins MH, Hubbell DH (1979). Plant growth substances produced by Azospirillum brasilense and their effect on the growth of pearl millet (Pennisetum americanum L.). Appl. Environ. Microbiol. 37:1016-1024. |
|
|
Travaglia C, Masciarelli O, Fortuna J, Marchetti G, Cardozo P, Lucero M, Zorza E, Luna V, Reinoso, H (2015). Towards sustainable maize production: Glyphosate detoxification by Azospirillum sp. and Pseudomonas sp. Crop Prot. 77:102-109. |
|
|
United States Department of Agriculture (USDA) (2017). World corn supply and use. World agricultural supply and demand estimates. |
|
|
United States Department of Agriculture (USDA) (2018). World corn supply and use. World agricultural supply and demand estimates. |
|
|
Vazquez GH, Silva MRR, Sousa JFS, Baroles RD (2012). Fontes de Azospirillum brasilense e doses de nitrogênio em cobertura na cultura do milho. In. 29 Congresso nacional de milho e sorgo. Campinas: Instituto Agronômico/Associação Brasileira de Milho e Sorgo. pp. 1639-1645. |
|
|
Verona DA, Duarte Junior JB, Rossol CD, Zoz T, Costa ACT (2010). Tratamento de Sementes de Milho com Zeavit®, Stimulate® e Inoculação com Azospirillum spp. In. Congresso Nacional de Milho e Sorgo, 18. Goiânia: Associação Brasileira de Milho e Sorgo. |
|
|
Yang J, Kloepper JW, Ryu CM (2009). Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 14:1-4. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


