ABSTRACT
Javanese ginseng (Talinum paniculatum) is an herb and one of the traditional medicinal plants in Indonesia which accumulate saponin in the root organ. However, slow root growth hampers the accumulation of the compound in this plant. Therefore, in vitro culture of adventitious roots offers an alternative way to increase the production of saponin. Furthermore, this study was performed to investigate the effect of methyl jasmonate (MeJA) and salicylic acid (SA) on the growth and saponin content in adventitious root of Javanese ginseng. Adventitious root culture was induced on solid Murashige and Skoog (MS) medium supplemented by 10 µM IBA (indole-3-butyric acid). Subsequently, adventitious roots were subcultured into a half strength MS liquid medium with the addition of different concentrations of MeJA or SA and incubated for 5, 10 and 15 days. Despite inhibition on the root growth, saponin production increased by 1.5 and 1.3 fold upon elicitation with 0.2 mM MeJA and SA for 15 days, respectively. Thus, MeJA and SA elicitations regulate saponin biosynthesis in adventitious root culture of Javanese ginseng as a time and dose-dependent manner.
Key words: Adventitious root, elicitation, Javanese ginseng, saponin, Talinum paniculatum.
Talinum paniculatum (family Talinaceae, is a succulent plant that grows up to 100-120 cm tall and characterized by a large and many-flowered terminal panicle with pink color. In Indonesia, this plant is also called Javanese ginseng as it has swelling roots similar to Panax ginseng root and its extract is used as a traditional medicine for multiple diseases (Manuhara et al., 2015). Investigation of root extract from Javanese ginseng resulted in the identification of triterpenoid saponin mixtures. Further characterization from the same genus resulted in the
identification of oleanane-type saponins which is responsible for different pharmacological activities such as estrogenic, antifertility, antibacterial, antifungal, antioxidant, and cytotoxic activities (Kohda et al., 1992; Reis et al., 2015; Thanamool et al., 2013).
In the natural habitat, the root of Javanese ginseng has a slow growth rate. Generally, it takes about 2-3 years to get more than 100 g of root per plant (Manuhara et al., 2015). Therefore, in vitro root culture is essential for preservation of this plant, especially to support sustainable saponin production. Adventitious root culture has been used to produce wide range of secondary metabolites which have been naturally accumulated in root organ. The advantages of adventitious root culture are the root, not influenced by geotropism, fast growth of root branch and genetically stable (Murthy et al., 2016; Inabuy et al., 2017).
Methyl jasmonate (MeJA) is a widely used elicitor to induce secondary metabolite compounds in plant cell culture. MeJA and its derivatives have been proposed to be the key signaling compounds in the process of elicitation leading to the accumulation of secondary metabolites (Ramirez-Estrada et al., 2016). Salicylic acid (SA) is another elicitor that is widely studied as a stressed molecule signal to respond to the pathogen in plants (Hernández et al., 2017). Furthermore, SA was reported to enhance phytoalexin production in cell culture and root culture of several medicinal plants such as Withania somnifera, Anisodus luridus and Lepechinia caulescens (Sivanandhan et al., 2013; Qin et al., 2014; Vergara Martínez et al., 2017). Among all biotic and abiotic elicitors, MeJA and SA are the most important elicitors used as inducers of triterpene saponin production (Moses et al., 2013). Therefore, this experiment was established to compare the effect of different elicitors (MeJA and SA) in various concentrations (0.05, 0.1 and 0.2 mM) and duration of elicitation (5, 10 and 15 days) on saponin production from adventitious roots culture of Javanese ginseng.
Adventitious root culture
Adventitious roots were induced from the shoot culture of Javanese ginseng grown on a phytohormone-free Murashige and Skoog (MS) solid medium (Murashige and Skoog, 1962)containing 30 g/L sucrose and 8 g/L agar. 20 mg of adventitious roots were then isolated and placed onto a half-strength MS solid medium supplemented with 1 µM and 10 µM of IBA (indole-3-butyric acid) (Duchefa Biochemie, the Netherlands) or NAA (1-napthaleneacetic acid) (Duchefa Biochemie, the Netherlands). Root cultures were incubated at 25±1°C in the dark. After 28 days, adventitious roots were harvested and the respective number of lateral roots that appear as well as final biomass were measured. Subsequently, the adventitious roots were subcultured in a 100 ml Erlenmeyer flask containing 15 ml of half-strength MS liquid medium for treatment with elicitors.
Effect of MeJA and SA on adventitious root growth and saponin production
Treatments of adventitious roots were performed with an inoculum of about 0.7 g fresh weight root segment in 20 ml of half-strength MS liquid medium supplemented with 10 µM IBA and 15 g/L sucrose. Different concentrations of MeJA (0, 0.05, 0.1, and 0.2 mM) and SA (0, 0.05, 0.1, and 0.2 mM) were added in the medium. The root was harvested after elicitation for 5, 10, and 15 days. After that, dry weight and saponin content were measured.
Extraction and determination of total saponin content
Roots were collected from MeJA and SA treated and non-treated samples at different incubation time. Subsequently, the dry weight was measured after drying the roots in the oven at 50°C for 24 h. All samples were stored on the desiccator until constant weight was reached.
Thirty milligram of dried sample was ground to a fine powder and dissolved in 1.5 ml of 96% methanol. The mixture was sonicated for saponin extraction using sonicator (Branson sonicator 3510) for 1 h and then centrifuged at 10,000 rpm for 10 min. Collected supernatant was used for total ginseng saponin analysis.
Total saponins were measured through spectrophotometer (Optima Japan SP3000 nano) as described by Fiallos-Jurado et al. (2016)with minor modification. The Liebermann-Burchard (LB) reagent was used for saponin quantification as it is capable of producing a light brown staining if this compound is present in a sample. The LB reagent consists of a 1:5 mixture of acetic acid and sulfuric acid, respectively. After mixing 1 ml sample solution with 3.5 ml of LB reagent, absorbance at 540 nm was measured in all samples after 20 min. A calibration curve based on oleanolic acid was used to determine the final concentration of saponin (mg/ml) in each solution. The total saponin content was calculated on the basis of dry weight.
Statistical analysis
Data were analyzed using one-way ANOVA followed by Duncan’s multiple range tests. All statistical analyses were performed at the level of P value less than 0.05 using SPSS 23.0 (SPSS Inc. USA).
Adventitious root cultivation
The number of lateral roots and biomass was affected by auxin type and respective concentration. In general, roots treated by IBA form whitish yellow lateral roots with numerous root hairs while those treated by NAA form brown yellowish lateral roots and also callus in their main roots. The roots formed in a medium containing NAA were also shorter and thicker than roots grown in a medium containing IBA.
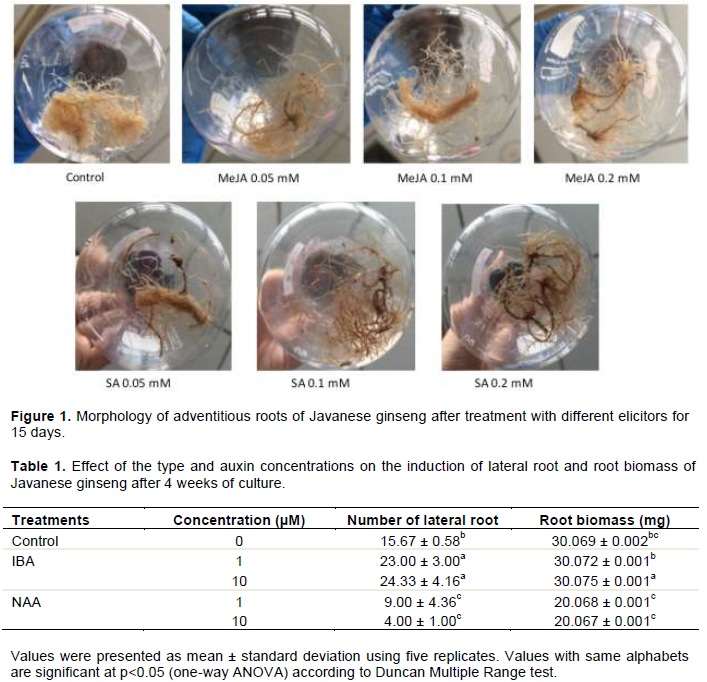
The number of lateral roots that emerged from adventitious root explant varied in all treatments (Figure 1). The result showed that IBA induced the development of more lateral roots compared with control, while NAA showed a contrasting result. Furthermore, there was no significant difference between the roots induced by 1 µM and 10 µM IBA in terms of the number of lateral roots, yet 10 µM IBA resulted in the highest root biomass (Table 1). The adventitious roots incubated in 10 µM IBA has growth rate of 0.035 g/day with doubling time for 27 days. Therefore, IBA 10 µM was further added to the medium for adventitious root cultivation.
Effect of MeJA and SA on adventitious root growth
Both treated and non-treated roots were growing on the liquid MS medium. However, the treated roots showed a slower growth rate compared to non-treated control. Furthermore, the color of treated roots changed and tends to be darker compared to that in control (Figure 1). Dry weight of adventitious root culture during the 5, 10, and 15 days after treatment with various concentrations of MeJA and SA are shown in Figure 2. In each incubation time, root biomass decreased from 32-37% when treated with MeJA. Similar result was also observed on SA treated samples in which root biomass decreased from 23 to 35%.
Effect of MeJA and SA elicitation on saponin production
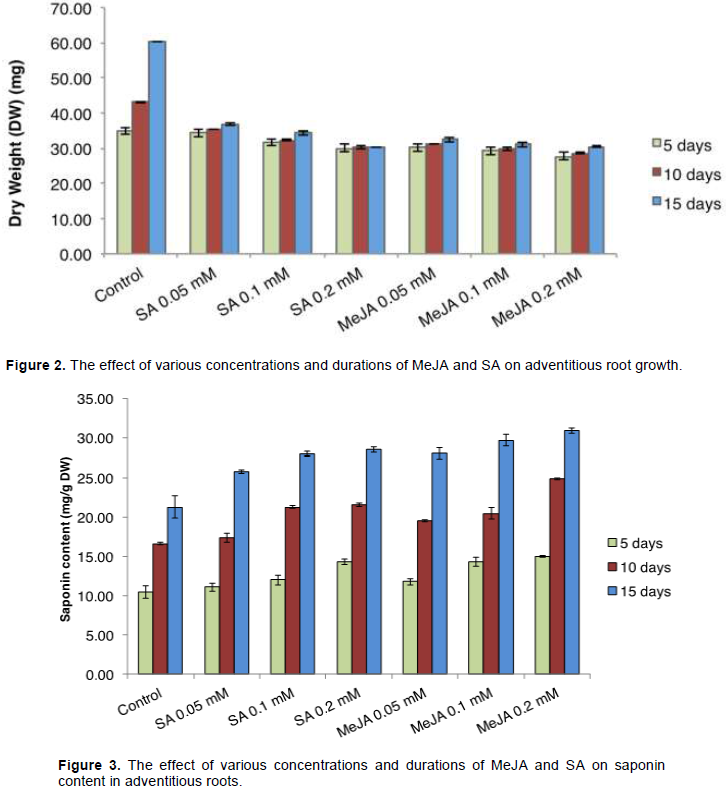
Increased concentration of MeJA and SA resulted in the accumulation of saponin content in the adventitious roots (Figure 3). The longer the incubation period which allow contact with elicitors, the more there is saponin accumulation in the root culture. At the 5-day treatment, saponin content increased as long as there is increase in elicitor concentration; however, the results slightly increase. This is because during the 5 days incubation, the roots are still in the adaptation stage with the new medium. At the 10-day treatment, saponin content at the root also increase with increasing elicitor concentration of 0.2 mM of MeJA and SA increased saponin content of about 1.4 and 1.2 times respectively compared to control.
Amount of total saponin increased with increasing MeJA and SA concentrations, and reached a maximum accumulation up to 30 mg/g dry weight (DW) upon treatment with 0.2 mM MeJA for 15 days. In every treatment compared to control, MeJA with concentration of 0.05, 0.1 and 0.2 mM increased saponin content by 1.2, 1.3 and 1.5 fold, respectively, whereas SA with concentration of 0.05, 0.1 and 0.2 mM increased saponin content by 1.1, 1.26 and 1.3 fold, respectively.
In vitro tissue cultures have been selected as alternatives for the production of desired saponin compounds in many plants (Espinosa-Leal et al., 2018; Leonard et al., 2018). These techniques could solve different problems associated with saponin extraction from natural-grown plants as well as offer a more sustainable mean for the production of this secondary compound. Furthermore, some plant secondary compounds are controlled in a tissue-specific manner, thus root or shoot culture are more preferred as the production vehicles (Faizal et al., 2013). Taken together, plant tissue culture could help to facilitate the improvement of saponin production through biotechnological approaches.
In this study, adventitious root culture was induced from Javanese ginseng by employing different concentrations of IBA and NAA. IBA was found to be more superior over NAA which may be due to the effects of IBA on the induction of lateral root. In morphological and histological aspects, the roots formed under IBA treatment developed normally compared to NAA. Based on the study conducted by Pacurar et al. (2014), growth regulators such as IBA have a higher potential to induce roots than other auxin hormones. The study also reported that IBA has a prominent effect on the stimulation of in vitro lateral roots from a mature root tissue. Similarly, adventitious roots of P. ginseng with IBA-containing medium were more effective in inducing lateral root and root growth compared to NAA (Murthy and Paek, 2016).
Many important plant secondary metabolites are hindered by low production titer which could prevent their commercial application. One of the technical approaches used to overcome this problem is application of elicitor to trigger a physiological response in plants and results in the accumulation of secondary metabolites. Moreover, elicitation depends on many factors such as elicitor type and concentration, the growth stage of the culture at the time of elicitor addition and incubation period with elicitor (Ramirez-Estrada et al., 2016). In this context, we used different concentrations of MeJA and SA together with different incubation periods to improve saponin production in adventitious roots of Javanese ginseng.
Both treatment of MeJA and SA induced a negative growth effect on adventitious roots of Javanese ginseng, which is a common phenomenon observed in plants. MeJA is suggested to inhibit root growth by interfering with transmembrane auxin flux pathway (Yan et al., 2016). Likewise, it has been reported that root growth in Arabidopsis was decreased upon modulation with SA signaling pathway (Rivas-San Vicente and Plasencia, 2011). Similarly, the elicitation using MeJA and SA on P. ginseng adventitious root culture showed inhibition of the root growth up to 25% (Ali et al., 2006)as reported in other plants (Saini et al., 2013; Eichmann and Schäfer, 2015).
The different growth shown by the treated roots indicate a stress condition. SA as a phenolic compound was produced naturally on every plant, and accumulation of phenolic compound on roots might occur as indicated by browning in response to additional SA put into the medium. In view of the inverse relationship between the production of biomass and the accumulation of the secondary metabolite, the root growth depression arising from elicitation may favor the synthesis of the secondary metabolite.
Amount of total saponin in the adventitious root cultures of Javanese ginseng increased with increasing MeJA or SA concentrations, and reached a maximum following the addition of 0.2 mM MeJA for 15 days to the cultivation medium. Previous study in whole root culture of G. glabra showed that 2 mM MeJA and 1 mM SA increased respective glycyrrhizin production by 3.8 and 4.5 fold after elicitation for 24 h (Shabani et al., 2009); whereas, adventitious root culture of P. ginseng required 7 days incubation with 0.2 mM of MeJA or SA for increasing the production of ginsenoside to 4 and 3 fold, respectively (Ali et al., 2006). The prolonged elicitation for 35 days in root culture of Centella asiatica significantly increased the accumulation of saponin by 4 to 6 fold (Mangas et al., 2008). The result indicated that the optimum concentration of signaling compounds varies according to the species. Also, contact time with elicitor was an important factor for enhanced production of secondary metabolite. In conclusion, MeJA and SA are potentially used to improve the yield of saponins in adventitious root cultures in Javanese ginseng.
The authors have not declared any conflict of interests.
This research was partially funded by the Ministry of Research, Technology and Higher Education, the Republic of Indonesia under the scheme of excellent research for university grant (Contract No. 3150/I1.C02.2/ KU/2018).
REFERENCES
|
Ali MB, Yu K-W, Hahn E-J, Paek K-Y (2006). Methyl jasmonate and salicylic acid elicitation induces ginsenosides accumulation, enzymatic and non-enzymatic antioxidant in suspension culture Panax ginseng roots in bioreactors. Plant Cell Reports 25(6):613-620.
Crossref
|
|
|
|
Eichmann R, Schäfer P (2015). Growth versus immunity-a redirection of the cell cycle? Current Opinion in Plant Biology 26:106-112.
Crossref
|
|
|
|
|
Espinosa-Leal CA, Puente-Garza CA, García-Lara S (2018). In vitro plant tissue culture: means for production of biological active compounds. Planta 248(1):1-18.
Crossref
|
|
|
|
|
Faizal A, Foubert K, Lambert E, Storme N, Claeys M, Apers S, Geelen D (2013). Saponin production is not qualitatively changed upon callus regeneration in the medicinal shrub Maesa perlarius. Plant Growth Regulation 70(1):39-48.
Crossref
|
|
|
|
|
Fiallos-Jurado J, Pollier J, Moses T, Arendt P, Barriga-Medina N, Morillo E, Arahana V, de Lourdes Torres M, Goossens A, Leon-Reyes A (2016). Saponin determination, expression analysis and functional characterization of saponin biosynthetic genes in Chenopodium quinoa leaves. Plant Science 250:188-197.
Crossref
|
|
|
|
|
Hernández JA, Diaz-Vivancos P, Barba-Espín G, Clemente-Moreno MJ (2017). On the role of salicylic acid in plant responses to environmental stresses. In: Nazar R, Iqbal N, Khan NA, eds. Salicylic Acid: A Multifaceted Hormone. Springer Singapore: Singapore pp. 17-34.
Crossref
|
|
|
|
|
Inabuy F, Fischedick JT, Lange I, Hartmann M, Srividya N, Parrish AN, Xu M, Peters RJ, Lange BM (2017). Biosynthesis of diterpenoids in Tripterygium adventitious root cultures. Plant Physiology 175(1):92-103
Crossref
|
|
|
|
|
Kohda H, Yamaoka Y, Morinaga S, Ishak M, Darise M (1992). Saponin from Talinum triangulare. Chemical and Pharmaceutical Bulletin 40:2557-2558.
Crossref
|
|
|
|
|
Leonard J, Seth B, Sahu BB, Singh VR, Patra N (2018). Statistical optimization for enhanced bacoside A production in plant cell cultures of Bacopa monnieri. Plant Cell, Tissue and Organ Culture 133(2):203-214.
Crossref
|
|
|
|
|
Mangas S, Moyano E, Osuna L, Cusido RM, Bonfill M, Palazon J (2008). Triterpenoid saponin content and the expression level of some related genes in calli of Centella asiatica. Biotechnology Letters 30(10):1853-1859.
Crossref
|
|
|
|
|
Manuhara YSW, Kristanti AN, Utami ESW (2015). Optimization of culture conditions of Talinum paniculatum Gaertn. adventitious roots in balloon type bubble bioreactor using aeration rate and initial inoculum density. Asian Journal of Biological Sciences 8:83-92.
Crossref
|
|
|
|
|
Moses T, Pollier J, Thevelein JM, Goossens A (2013). Bioengineering of plant (tri)terpenoids: from metabolic engineering of plants to synthetic biology in vivo and in vitro. New Phytologist 200(1):27-43.
Crossref
|
|
|
|
|
Murashige T, Skoog F (1962). A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiologia Plantarum 15:473-497.
Crossref
|
|
|
|
|
Murthy HN, Dandin VS, Paek K-Y (2016). Tools for biotechnological production of useful phytochemicals from adventitious root cultures. Phytochemistry Reviews 15(1):129-145.
Crossref
|
|
|
|
|
Murthy HN, Paek KY (2016). Panax ginseng adventitious root suspension culture: Protocol for biomass production and analysis of ginsenosides by high pressure liquid chromatography. In: Jain SM, ed. Protocols for In Vitro Cultures and Secondary Metabolite Analysis of Aromatic and Medicinal Plants, Second Edition. Springer New York: New York, NY, pp. 125-139.
|
|
|
|
|
Pacurar DI, Perrone I, Bellini C (2014). Auxin is a central player in the hormone cross-talks that control adventitious rooting. Physiologia Plantarum 151(1):83-96.
Crossref
|
|
|
|
|
Qin B, Ma L, Wang Y, Chen M, Lan X, Wu N, Liao Z (2014). Effects of acetylsalicylic acid and UV-B on gene expression and tropane alkaloid biosynthesis in hairy root cultures of Anisodus luridus. Plant Cell, Tissue and Organ Culture 117(3):483-490.
Crossref
|
|
|
|
|
Ramirez-Estrada K, Vidal-Limon H, Hidalgo D, Moyano E, Golenioswki M, Cusidó RM, Palazon J (2016). Elicitation, an effective strategy for the biotechnological production of bioactive high-added value compounds in plant cell factories. Molecules 21(2):182.
Crossref
|
|
|
|
|
Reis LFCD, Cerdeira CD, DE PBF, Silva JJd, Coelho LFL, Silva MA, Marques VBB, Chavasco JK, Alves-Da-Silva G (2015). Chemical characterization and evaluation of antibacterial, antifungal, antimycobaterial, and cytotoxic activities of Talinum Paniculatum. Revista do Instituto de Medicina Tropical de São Paulo 57:397-405.
Crossref
|
|
|
|
|
Rivas-San Vicente M, Plasencia J (2011). Salicylic acid beyond defence: its role in plant growth and development. Journal of Experimental Botany 62(10):3321-3338.
Crossref
|
|
|
|
|
Saini S, Sharma I, Kaur N, Pati PK (2013). Auxin: a master regulator in plant root development. Plant Cell Reports 32(6):741-757.
Crossref
|
|
|
|
|
Shabani L, Ehsanpour AA, Asghari G, Emami J (2009). Glycyrrhizin production by in vitro cultured Glycyrrhiza glabra elicited by methyl jasmonate and salicylic acid. Russian Journal of Plant Physiology 56(5):621-626.
Crossref
|
|
|
|
|
Sivanandhan G, Kapil Dev G, Jeyaraj M, Rajesh M, Arjunan A, Muthuselvam M, Manickavasagam M, Selvaraj N, Ganapathi A (2013). Increased production of withanolide A, withanone, and withaferin A in hairy root cultures of Withania somnifera (L.) Dunal elicited with methyl jasmonate and salicylic acid. Plant Cell, Tissue and Organ Culture 114(1):121-129.
Crossref
|
|
|
|
|
Thanamool C, Papirom P, Chanlun S, Kupittayanant S (2013). Talinum paniculatum (Jacq.) Gertn: a medicinal plant with potential estrogenic activity in ovariectomized rats. Internationl Journal of Pharmacy and Pharmaceutical Science 5:478-485.
|
|
|
|
|
Vergara Martínez VM, Estrada-Soto SE, Arellano-García JdJ, Rivera-Leyva JC, Castillo-Espa-a P, Flores AF, Cardoso-Taketa AT, Perea-Arango I (2017). Methyl jasmonate and salicylic acid enhanced the production of ursolic and oleanolic acid in callus cultures of Lepechinia caulescens. Pharmacognosy Magazine 13:S886-S889.
|
|
|
|
|
Yan S, Zhang T, Dong S, McLamore ES, Wang N, Shan X, Shen Y, Wan Y (2016). MeJA affects root growth by modulation of transmembrane auxin flux in the transition zone. Journal of Plant Growth Regulation 35(1):256-265.
Crossref
|
|