Full Length Research Paper

ABSTRACT
Information on microbial production of cytokinins and their effect on plant growth are limited. Therefore, the objective of the present study was to investigate cytokinins production by Pseudomonas sp., a major component of rhizobacteria with multiform and diverse activities, which enhance plant growth by synthesizing various secondary metabolites. In the present investigations, thirty Pseudomonas isolates were isolated from the rhizosphere of Pyrus and Malus and were screened for cytokinins production (50 to 210 μg/ml). Four strains viz PN-4-SAN, PN-10-SAN, AN-2-NAG and AN-4-NAG were selected on the basis of their higher cytokinins production. The maximum cytokinins production was observed at 72h incubation period in nutrient broth at pH 7.0 under shaken condition at 28°C. Cytokinins were extracted, purified and evaluated by thin layer chromatography and specific bioassay method.
Key words: Cytokinins, plant growth promoting rhizobacteria (PGPR), Pseudomonas sp., thin layer chromatography, radish cotyledon bioassay.
INTRODUCTION
Fluorescent Pseudomonas species have emerged as largest and potentially most promising group of plant growth promoting rhizobacteria (PGPR). Their potential to synthesize different secondary metabolites like phytohormones with diverse and multiform activities is an important function to improve plant growth and health (Kloepper et al. 1988). Phytostimulatory effect of PGPR may be initiated by several ways but the production of phytohormone such as cytokinins and facilitating resource acquisition by such bacteria is direct mechanism of plant growth promotion (Garcia de Salamone et al., 2001; Glick, 2012; Ramadan et al., 2016) while indirect mechanism of action may involve antibiotic production, depletion of iron from the rhizosphere, synthesis of antifungal metabolites, production of fungal cell wall lysing enzymes, competition for sites on roots and induced systemic resistance (Glick, 2012). Phytohormone, cytokinins can be produced by many bacterial genera and its occurrence was observed in many terrestrial and rhizospheric soil (Kampert and Strzelcyk, 1984; Arkhipova et al., 2007; Hussain and Hasnain, 2009) which suggested that bacterial cytokinins may affect the growth and development of plants. Recent studies confirmed that, a number of bacterial species mostly associated with the plant rhizosphere, are found to be associated with cytokinins production including Flavobacterium, Acinetobacter, Bacillus, Arthrobacter, Aerobacter, Azospirillum and Pseudomonas sp. (Maruyama et al., 1986; Perig et al., 2007; Arkhipova et al., 2007; Ortiz-Castro et al., 2008). Cytokinins have key regulatory roles in plant growth and development. They promote seed germination, de novo bud formation, release of buds from apical dominance, stimulation of leaf expansion and of reproductive development besides on, retardation of senescence (Mok, 1994). Some of these effects have been observed in cucumber cotyledon when inoculated with Pseudomonas aeruginosa E2 (Hussain and Husnain, 2009).
Cytokinins influence cell division activity in embryonic as well as mature plants by altering the size and activity of meristems (Werner et al., 2001). Yang et al. (2002) showed that the rate of endosperm cell division is closely associated with cytokinin level in endosperm. They also reported that exogenous kinetin significantly increase the number of endosperm cells and grain weight. Morris and Powell (1987) first reported on genes, specifying cytokinins biosynthesis from phytopathogenic bacteria notably Agrobacterium tumefaciens (tmr, tzs) and Pseudomonas syringae pv, savastanoi (ptz). Genes encode prenyl transferase is responsible for cytokinins biosynthesis. High levels of cytokinins were found toincrease the resistance of plants to some viral pathogens and herbivores (Ballare, 2011).
Großkinsky et al. (2016) identified the ability of Pseudomonas fluorescens G20-18 to efficiently control P. syringae infection in Arabidopsis and cytokinin production which was identified as a key determinant for this biocontrol effect on the hemibiotrophic bacterial pathogen, allowing maintenance of tissue integrity and ultimately biomass yield. Naseem and Dandekar (2012) also reported that cytokinin confers protection to Arabidopsis against infection with Pseudomonas syringae pv. tomato (Pto).
However, to date little information exists on the diversity of plant growth promoting rhizobacteria associated with apple and pear trees. Moreover, in apple and pear growing region of Himachal Pradesh, cytokinins producing Pseudomonas population has never been estimated.
In present work, plant growth regulators producing Pseudomonas sp. from rhizospheric soil of apple and pear trees were isolated and characterized for production of one of the important plant growth regulators i.e. cytokinins.
MATERIALS AND METHODS
Isolation of fluorescent Pseudomonas species from rhizosphere of apple and pear trees
Soil sampling
Soil samples were collected from rhizosphere of ten years old orchards of Pyrus (pear) from Sanyardi (31°41’42.1” N, 76°55’56.7” E) and Malus (apple) from Nagwain (31°48’57.9” N, 77°10’46.1” E) regions of Mandi district (Himachal Pradesh), India during autumn season (25°C). The pH of rhizospheric soil of pear and apple was found to be 7.3 and 7.5, respectively and electrical conductivity is 195 and 210 μS/cm, respectively. Four samples were taken around few to 10 cm apart from roots and 30 to 45 cm deep from five different plants and mixed to make it one composite sample.
Isolation and Identification
Total bacterial count viz- a-viz fluorescent Pseudomonas sp. was isolated on Nutrient agar and King’s B media (Hi-media), respectively from soil samples. The proposed isolates were characterized on the basis of morphological, biochemical and physiological analyses as prescribed in Bergey’s manual of systematic bacteriology by Kreig and Holf (1984).
Screening of isolates for cytokinins production
For production of cytokinins, Pseudomonas strains (Inoculum size 108 cfu/ml) were grown in nutrient broth for 72h at 28 ± 2°C under shaken conditions. Supernatant was prepared by centrifugation of cultures at 10,000 rpm for 20 min and was stored in deep fridge or at 4°C. Radish cotyledons expansion test was employed (Letham, 1971) for assay of cytokinins like, substances; the radish seeds (Raphanus sativus L. cultivars Japanese white) which were germinated in total darkness for 48 h at 28°C. After removing the seed coat, smaller cotyledons were transferred to sterilized Petri dishes containing the test solution, water (as blank) and standard solution (kinetin) on filter paper strips. Twelve cotyledons were placed in each Petri dish and kept at 25°C under fluorescent light for 3 days. Then cotyledon on filter paper strips in Petri dish were blotted, dried and weighed. The bioassay response (final weight-initial weight) was expressed as an increase in weight. Concentration of cytokinins present in the extract was calculated by preparing standard curve using kinetin as standard (100 to 1000 µg/ml).
Effect of culture conditions on production of cytokinins by Pseudomonas sp.
The test organisms (107 cfu) were inoculated in five different media; succinate media, king’s media, nutrient media, peptone water and trypticase soyabroth. Flasks were incubated at 28°C for 24, 48, 72 h under shaker at 90 rpm. After this step, supernatant were picked up by centrifugation in multifuge XIR (Thermoscientific Germany) at 10,000 rpm for 30 min at 4°C and were further used for estimation of cytokinins.
Extraction and separation of cytokinins
Cytokinins like substance were extracted by the method of Tien et al. (1979) with slight modifications. Acidified supernatant extracted with di-ethyl ether and partitioned with n- butanol. Residue was dissolved in 0.1 N HCl (10 ml) and passed through Dowex-50 column chromatography and eluted with sodium hydroxide. Extracted and concentrated fraction was diluted in distilled water. Collected fraction was separated by thin layer chromatography by spotted (100 µl) on silica gel-G plates and developed in n-butanol: ammonium hydroxide (I N): water (7:1:2). The plates were examined under U.V light (254 nm) and observed for the fluorescent spots.

Evaluation of cytokinins by Radish cotyledons expansion test
The partially purified cytokinins were evaluated by radish cotyledon expansion test. Radish seeds were surface sterilized with 0.1% HgCl2 for 3 min and rinsed five times with sterile distilled water and germinated in total darkness for 48 h at 28°C. After removing the seed coat smaller cotyledons were transferred to sterilize Petri dishes containing 7ml test solution (extracted sample), water (control) and standard solution (Kinetin) 100 to 1000 µg on filter paper strips. Twelve cotyledons were placed in each Petri dish and were incubated at 25°C under fluorescent light for three days. Each treatment was replicated three times. The cotyledons were filtered and weighed. The bioassay doses response curve drawn (initial weight to final weight) was expressed as increase in weight against concentration of cytokinins present in the extract and calculated by preparing standard doses responses curve by using kinetin as standard (Letham, 1971).
Statistical analysis
The data obtained was subjected to analysis of variance technique using completely randomized design (CRD).
RESULTS
The rhizospheric soil samples were evaluated for the status of total bacterial population viz a viz Pseudomans sp. Total bacterial count of rhizospheric soil of apple and pear was 6.1 and 6.3 log cfu/g of soil, respectively. A total of seventeen Pseudomonas isolates were obtained from rhizosphere soil of pear plant and thirteen were obtained from rhizosphere soil of apple plant up to its maturity level. On the basis of morphology and biochemical analyses, these isolates were identified as Gram negative, rod shaped bacteria which produce fluorescence under ultra violet radiations and showed positive test for catalase and oxidase.
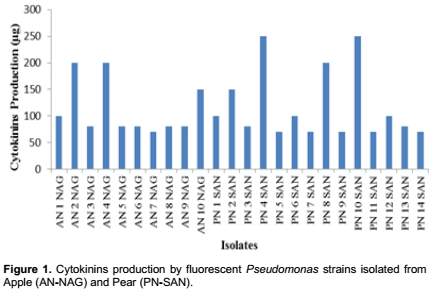
In our studies it was found that all Pseudomonas isolates produced cytokinins in the range of 70 to 250µg/ml (Figure 1). Four strains viz PN-4-SAN, PN-10-SAN, AN-2-NAG and AN-4-NAG were selected on the basis of their higher cytokinins production. The efficient isolates produced cytokinin like substances in the stationary growth phase i.e. at 72h of incubation period at 28°C in nutrient broth (Table 1).

The homogeneity of the partially purified cytokinins along with kinetin as standard was checked by thin layer chromatography. Cytokinins gave the maximum Rf value of 0.75. Brown spots corresponding to, cytokinins or cytokinins like substances were visible when sprayed with reagent (Table 2 and Figure 2). As the result showed partially purified cytokinins extracted samples from Pseudomonas sp. PN-4-SAN, PN-10-SAN, AN-2-NAG and AN-4-NAG have increased the weight of radish cotyledons by 0.02 to 0.2 cm respectively that correspondent to 100 to 200 µg/ml as calculated from the dosage response curve of kinetin (Table 3 and Figure 3).




DISCUSSION
Cytokinins are important class of phytohormones which are adenine derivatives. Phytostimulatory effects of PGPR may be initiated by several ways but cytokinin production by such bacteria is the direct mechanism to improve plant growth (Ortiz-Castro et al., 2008; Remans et al., 2008). Cytokinins producing bacteria i.e. Pseudomonas, Rhizobium, Bacillus and Agrobacterium have been isolated from various crop plants including maize, wheat, tomato, sugarcane, mustard, carrot and sunflower (Hussain and Hasnain, 2009). However, in our studies, Pseudomonas strains producing cytokinins were obtained from rhizospheric soil of apple and pear. Significant difference was observed in total bacterial and Pseudomonas communities of apple and pear orchards.
In this study, cotyledon bioassay was used to identify efficient Pseudomonas strains producing cytokinins like substance, as it was already found to be convenient and sensitive by providing a more linear response (Dumbroff and Brown 1976). Based on the data presented, the cytokinins yield was maximized (280 µg/ml) at 72 h of incubation at pH7 under shaken conditions at 28°C. It might be due to the reason that cultural conditions play an important role in cellular growth and biological activities of microorganisms as stated by Bentley and Keil (1962). Hussain and Hasnain (2009) reported cytokinin as equivalent to 1091.9 ng ml-1 in the late stationary phase culture of Bacillus licheniformis. In another study, Taller and Wong (1989) determined cytokinins as equivalent to 0.75 μg of kinetin per litre in Azotobacter vinelandii culture medium while Karnwal and Kaushik (2011) reported 5.5 and 2.9 pmol/ml of cytokinins per litre for P. fluorescens and Pseudomonas aeruginosa, respectively. However, Lichter et al. (1995) analyzed different forms of cytokinins that is, zeatin, zeatin riboside, isopentenyladenin and two immunoreactive zeatin type compounds in the culture supernatant of pathogenic strain Erwinia herbicola and quantified zeatin in range of 200 to 300 ng/ml and zeatin riboside 13 to 107 ng/ml. So the selected Pseudomonas isolates were found to be quite efficient for the production of cytokinins like substances.
We have checked partially purified cytokinins like substance by thin layer chromatography having Rf value 0.72 to 0.75 (Table 2 and Figure 2) and radish cotyledon test (Table 3 and Figure 3). Hussain and Hasnain (2009) studied the production of cytokinins like substances using chromatography and bioassays methods and however, they found highest biological activity exhibited in substances located at the Rf 0.5, 0.54 and 0.58 in Bacillus licheniformis, Bacillus subtilis and P. aeruginosa as cytokinins production was found to be less in comparison of our findings. These results, therefore, indicate that Pseudomonas sp. can be used as a potent bacterial source to obtain high yields and provide a good alternative for cytokinins biosynthesis. As compared to cytokinins obtained from other alternatives, they are less expensive and less time consuming. So, in future these be used for the commercial production of cytokinins.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Arkhipova TN, Prinsen E, Veselov SU, Martinenko EV, Melentiev AI, Kudoyarova GR (2007). Cytokinin producing bacteria enhance plant growth in drying soil. Plant Soil. 292:305-315. |
|
|
Ballare CL (2011). Jasmonate-induced defenses: a tale of intelligence, collaborators and rascals. Trends Plant Sci. 5:249-257. |
|
|
Bentley R, Keil J G (1962). Tetronic acid biosynthesis in molds. II. Formation of penicillic acid in Penicillium cyclopium. J. Biol. chem. 237:867-873. |
|
|
Dumbroff EB, DCW Brown (1976). Cytokinin and inhibitor activity in roots and stems of sugar maple seedlings through the dormant season. Can. J. Bot. 54:191-197. |
|
|
Garcia de Salamone IE, Hynes RK, Nelson LM (2001). Cytokinin production by plant growth promoting rhizobacteria and selected mutants. Can. J. Microbol. 47:404-411. |
|
|
Glick BR (2012). Plant Growth-Promoting Bacteria: Mechanisms and Applications. Scientifica. Article ID 963401, 15 pages. |
|
|
Großkinsky DK, Tafner R, Moreno MV, Stenglein SA, García de Salamone IE, Nelson LM, Novák O, Strnad M, van der Graaff E, Roitsch T (2016). Cytokinin production by Pseudomonas fluorescens G20-18 determines biocontrol activity against Pseudomonas syringae in Arabidopsis. Sci. Rep. 6:23310. |
|
|
Hussain A, Hasnain S (2009). Cytokinin production by some bacteria: Its impact on cell division in cucumber cotyledons. Afr. J. Microbiol. Res. 3:704-712. |
|
|
Kampert M, Strzelczyk E (1984). Effect of pH on production of cytokinin-like substances by bacteria isolated from soil, rhizosphere and mycorrhizosphere of pine (Pinus silvestris L.). Acta. Microbiol. Pol. 33:77-85. |
|
|
Karnwal A, Kaushik P (2011). Cytokinin production by fluorescent Pseudomonas in the presence of rice root exudates. Arch. Phytopathol. Plant Protect. 44:1728-1735. |
|
|
Kreig NR, Holf JG (1984). Bergeys Manual of Systematic Bacteriology. William and Wilkins, Baltimore, USA. |
|
|
Letham A (1971). Regulator of cell division in plant tissues XII. A cytokinin bioassay using excised radish cotyledons. Physiol. Plant. 25:391-396. |
|
|
Lichter A, Manulis S, Sagee O, Gafni Y, Gray J, Meilan R, Morris RO, Barash I (1995). Production of cytokinins by Erwinia herbicola pv. gypsophilae and isolation of a locus conferring cytokinin biosynthesis. Am. Phytopathol. Soc. 8:114-121. |
|
|
Maruyama A, Maeda M, Shimizu U (1986). Occurrence of plant hormone (cytokinin)-producing bacteria in the sea. J. Appl. Bacteriol. 61:569-574. |
|
|
Mok MC (1994). Cytokinins and plant development - an overview. In: Mok, D.W.S., Mok, M.C. (Eds.), Cytokinins: Chemistry, Activity and Function. CRC Press, New York, pp. 115-166. |
|
|
Morris RO, Powell GK (1987). Genes specifying cytokinins biosynthesis in prokaryotes. BioEssays 6:23-28. |
|
|
Naseem M, Dandekar T (2012). The Role of Auxin-Cytokinin Antagonism in Plant-Pathogen Interactions. PLoS Pathog. 8(11):e1003026. |
|
|
Ortiz-Castro R, Valencia-Cantero E, Lopez-Bucio J (2008). Plant growth promotion by Bacillus megaterium involves cytokinin signaling. Plant Signal. Behav. 3:263-265. |
|
|
Perrig D, Boiero ML, Masciarelli OA, Penna C, Ruiz OA, Cassan FD, Luna MV (2007). Plant-growth-promoting compounds produced by two agronomically important strains ofAzospirillum brasilense, and implications for inoculant formulation. Appl. Microbiol. Biotechnol. 75: 1143-1150. |
|
|
Ramadan EM, AbdelHafez AA, Hassan EA, Saber FM (2016). Plant growth promoting rhizobacteria and their potential for biocontrol of phytopathogens. Afr. J. Microbiol. Res. 10:486-504. |
|
|
Remans R, Beebe S, Blair M, Manrique G, Tovar E, Rao I, Croonenborghs A, Torres-Gutierrez R, El-Howeity M, Michiels J, Vanderleyden J (2008). Physiological and genetic analysis of root responsiveness to auxin-producing plant growth-promoting bacteria in common bean (Phaseolus vulgaris L.). Plant Soil. 302:149-161. |
|
|
Taller BJ, Wong TY (1989). Cytokinins in Azotobacter vinelandii culture medium. Appl. Environ. Microbiol. 55:266-267. |
|
|
Tien T M, Gaskin S, Hubbell D H (1979). Plant growth substances produced by Azospirillum brasilense and their effect on the growth of pearl millet (Pennisetum americanum L.). Appl. Environ. Microbiol. 37:1016-1024. |
|
|
Werner T, Motyka V, Strnad M, Schmulling T (2001). Regulation of plant growth by cytokinin. Proc. Natl. Acad. Sci. USA. 98:10478-10492 |
|
|
Yang J, Zhang J, Huang Z, Wang Z, Zhu Q, Liu L (2002). Correlation of cytokinin levels in the endosperm and roots with cell number and cell division activity during endosperm development in Rice. Ann. Bot. 90:369-377. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


