ABSTRACT
This study was aimed at determining the occurrence of Staphylococcus species in ready-to-eat (RTE) Rhynchophorus phoenicis (edible larvae) and Archachatina marginata (land snail) vended along Port Harcourt-Bayelsa route. Eighty samples from four locations were analysed employing standard techniques for proximate and microbiological analyses; virulence determination and antibiotic susceptibility. Data were statistically analyzed using ANOVA and t-test. The mean proximate results revealed the presence of protein (26.01/13.6%), lipid (18.9/3.88%), fibre (5.12/2.01%), ash (3.40/1.11%), moisture (13.47/59.5%) and carbohydrate (32.43/20.07%) for edible larvae/snail, respectively. Of the 80 samples examined, 33 (41.25%) and 52 (65.00%) had total viable bacterial counts and total staphylococcal counts above acceptable microbiological limits, respectively for RTE foods. Seven of the 81 characterized Staphylococcus produced the expected band of 950 bp with sea virulent genes while three produced expected bands of 550 bp with seb virulent genes. Three Staphylococcus aureus strains from edible larvae harboured both virulent genes. The virulent genes bearing Staphylococcus were 100% resistant to augmentin, ceftazidime and cloxacillin but showed varying resistance against ceftriaxone (57.14%), cefuroxime (28.57%), vancomycin (42.86%), oxacillin (42.86%) and cefoxitin (42.86%). The study showed that these RTE foods are potential sources of staphylococcal food poisoning in commuters; hence, food vendors need to conform to standard practice.
Key words: Edible larvae, land snail, staphylococcal enterotoxin A (sea), virulence.
Ready-to-eat foods (RTE) may be raw, minimally cooked or cooked foods and are vended for instant consumption without additional preparation. The usage of RTE foods prepared and sold by the street vendors has expanded in several African countries, including Nigeria (Makelele et al., 2015). There are varieties in the form of snacks, meats, salads, fruits and also beverages. Ready-to-eat foods have gained a lot of popularity such that they are available in the markets, roadsides and restaurants. Food vendor services is still on the increase, emerging to a new form referred to as migratory food vending especially on highways where RTE foods are highly exposed to microbial hazards. The pervasive routine of open presentation and exposure of ready-to-eat foods and the deficiencies of the existing food safety frameworks pose danger to the general wellbeing of commuters who patronize these foods. In Nigeria, African palm tree weevil larva (Rhyncophorus phoenicis) and African land snail meat (mainly Archachatina marginata) are regular edible insects and mollusks, respectively that are vended by migratory food vendors on the busy roads of Niger Delta states like Bayelsa, Delta, Edo, Imo, Rivers and also upper Cross-River Basins (Arene et al.,1999; Ekrakene and Igeleke, 2007). African palm tree weevil larva is popularly called edible larvae and it is widely recognized and prevalent in Tropical Africa (Tambe et al., 2013). It is principally sufficient in fat (66.61%) and protein (21.06%) (Womeni et al., 2012). It is a cherished delicacy not only in Southern Nigeria but other African countries like Cameroon. In ethnic/local languages, it is called ‘eruru’ in Ibo land, ‘orhu’ in Edo, ‘odon’ in Delta and ‘awon’ or ‘ekuku’ in Yoruba land. African palm weevil can be consumed raw, boiled, fried or roasted. Unlike other kinds of edible insects, African palm weevils are available year-round in variable quantities and also commonly seen in the southern parts of Nigeria rainforest zone while in Cameroon it is ubiquitous in moist swampy forest and Savannah zones (Fogoh et al., 2015). African land snails are domicile extensively in African countries like Nigeria, Ghana and also Central and Southern Africa where the climatic conditions are appropriate for rapid reproduction (Herbert and Kilburn, 2004). The two different land snails; A. marginata and Achatina achatina are well known edible snail groups in bounty in these localities (Ajayi et al., 2009). A report from USDA (2006) describes snail meat as top-notch sustenance, which is adequate in protein, lesser lipids and a decent wellspring of iron. Hence, these foods are patronized by travellers regardless of how they were processed by the migratory food vendors because of their nutritive properties. However, these are vehicles of transferring pathogens of general health significance to consumers since they may be inadequately processed or poorly handled during packaging and at point of sales. Although no reports of food borne illnesses have been linked with raw and roasted R. phoenicis larvae; they are nevertheless associated with a number of microorganisms, namely: Staphylococcus aureus, Bacillus species, Proteus species, Lactobacillus plantarum, Pseudomonas species, Escherichia coli, Enterococcus species, Proteus vulgaris, Serratia species, Acinetobacter species, Enterobacter species, Micrococcus species, Salmonella species, Listeria species, Aspergillus species, Penicillium species, Cladosporium species, Fusarium species, Rhizopus species and Mucor species (Wachukwu et al., 2002; Ekrakene and Igeleke, 2007; Oranusi and Braide, 2012; Amadi et al., 2014; Ebenebe and Okpoko, 2015; Daniel and Onilude, 2017).
Land snails have close association with microorganisms living in the soil environment whether wild or cultured, this affinity with snail and microorganisms is due to habitat filth, sewage and rotten materials (Agbonlahor et al., 1994). The microbial biota of snail in their unprocessed state is a contribution of their environment microbial community. Snails living in regular habitat can harbour pathogenic Enterobacteriaceae; at least 40 genera are listed including Salmonella, Proteus, Serratia, Enterobacter, Citrobacter, Pseudomonas and Klebsiella spp. (Fagbuaro et al., 2006). A number of authors have reported the occurrence of E. coli, Klebsiella spp., Salmonella spp., Enterobacter spp., Shigella spp., Citrobacter spp., Pseudomonas spp., Staphylococcus spp., Micrococcus spp., Proteus spp., Bacillus subtilis, Bacillus cereus, Lactobacillus spp. and Listeria monocytogenes in African land snail species (Adegoke et al., 2010; Nwuzo et al., 2016; Nyoagbe et al., 2016).
Staphylococcal food poisoning (SFP), caused by enterotoxin-producing S. aureus strains is an important foodborne illness in some countries (Wieneke et al., 1993; Le Loir et al., 2003). S. aureus are linked to many clinical cases (Adegoke and Komolafe, 2009). Thomas et al. (2007) described 19 types of staphylococcal enterotoxins (SEs); Group 1, classical emetic toxins: staphylococcal enterotoxin A (SEA), staphylococcal enterotoxin B (SEB), staphylococcal enterotoxin C1 (SEC1), staphylococcal enterotoxin C boy (SECbov), staphylococcal enterotoxin D (SED) and staphylococcal enterotoxin E (SEE), responsible for 95% food poisoning in man.
It is common to hear persons who changed locations complaining of stomach upset not usually linked to ready-to-eat foods consumed along the route because of varying incubation periods of food borne pathogens. The study was aimed at determining the occurrence of Staphylococcus spp. in ready-to-eat edible larvae (R. phoenicis) and land snail meats (A. marginata) vended along Port Harcourt-Bayelsa route.
Sample collection
A total of eighty ready-to-eat food samples comprising forty roasted edible larvae and forty land snail meats were purchased at different locations [Edible larvae (Mbiama, Tombia, Ekeki and Opokuma) and snail meats (Mbiama, Choba-Emohua, Ahoada and Opokuma)]. These ready-to eat foods were partially exposed and poorly packaged (Figures 1 and 2). The samples were aseptically stored in well labelled and sealed sterile transparent bags and transported immediately to the Microbiology laboratory for analyses.
Proximate analyses of the RTE food samples
The proximate parameters: ash content, moisture content, crude protein, crude lipid, crude fibre and total carbohydrate of the ready-to-eat samples were determined as described by Association of Official Analytical Chemists (AOAC, 2000). Crude protein was analysed using Kjedahl method, adopted from the procedures of James (1995) and Chang (2003). The percentage of the total nitrogen derived was converted to crude protein by multiplying by 6.25 (that is N × 6.25). Crude fat was performed by extraction with petroleum ether solvent. The defatted samples were analysed for ash content. Carbohydrate content of the food was determined by percentage difference in sum of other parameters (ash content, crude protein, crude fibre, and crude fat and moisture contents) from 100. Each parameter was conducted in duplicates.
Microbiological analyses
Sample preparation
Twenty-five grams of the food sample was transferred to the sterile blender (Philip, HR2001, China) macerated/diluted with 225 ml sterile peptone water (Oxoid, Basingstoke, UK) for 2 min. Serial dilution was done on the homogenized stock solution up to 10-6 as described by Makelele et al. (2015).
Determination of total viable counts
Aliquot (0.1 mL) of each of the selected sample dilutions was aseptically spread plated on the sterile solidified nutrient agar (NA: TM, Rajasthan, India) in triplicate. The inoculum was allowed to be absorbed into the agar plate, inverted and incubated at 30°C for 24 h. After incubation, average number of colony counts of the same dilution was recorded and colony forming units per gram (CFU/g) obtained and expressed as logarithm (log10 CFU/g) (Harrigan and Mc-Cance, 1990; Cheesbrough, 2010).
Enumeration and isolation of S. aureus
Aliquot (0.1 mL) of the selected sample dilutions were inoculated on freshly prepared, dry surface mannitol salt agar (MSA; Oxoid, Basingstoke, UK) and spread carefully on the entire surface of the agar plates. The inocula were allowed to absorb, thereafter, incubated at 30±2°C for 48 h. A colour change in the agar from red to yellow (fermentation of mannitol) indicated presumptive S. aureus colonies. Hence, mannitol positive colonies surrounded by yellow halo were counted (Makelele et al., 2015). Staphylococcus discrete colonies were counted, recorded and used to obtain staphylococci colony forming unit per gram and expressed in logarithm log10 CFU/g (Lancette and Benett, 2001).
Molecular characterization of isolated S. aureus
DNA extraction
The extraction of DNA was by boiling method described by Eruteya et al. (2014) and Vendruscolo et al. (2017). For DNA extraction, aliquot of 0.5 mL of 5 mL overnight Luria Bertani (LB) broth culture of bacterial isolates was transferred to 2 mL Eppendorf tubes. The cells were centrifuged (Dupont Sorvall MC-12V, biomedical division, USA) at 16099 g. for 3 min after addition of sterile distilled water. The supernatant was discarded while the harvested cells in the residue were vortexed (Vortexer VWR G-560, Scientific Industries, USA) and re-centrifuged after re-suspension in 500 µL of normal saline. DNA elution buffer (500 µL) was then added to the residues after discarding the supernatants and spun again. Thereafter, 300 µL DNA elution buffer was added to the residues and vortexed for about 3 min, and then heated with a heating block (Wealtec HB-2, Wealtec Corp, USA) at 95oC for 20 min. The heated bacterial suspensions were fast cooled on ice to -20°C for 10 min and spun for 3 min at 16099 g. The solutions containing the extracted genomic DNA (200 µL) were moved to 1.5 mL micro-centrifuge tubes and kept at Ë—20°C in the deep freezer for further downstream reactions.
The extracted DNA of the isolates were quantified using the Nanodrop 1000 spectrophotometer (Thermo Fisher Scientific, Inqaba Biotechnological Industries Ltd Pretoria, SA) which was used to measure the DNA concentration in ng/µL.
Amplification of sea genes for S. aureus strains
The sea genes from the suspected enterotoxigenic S. aureus isolates were amplified using the sea primers (Table 1) on DNA thermal cycler (GeneAmp PCR system) ABI 9700 Applied Biosystems, SA) at final volume of 30 µl for 35 cycles. The PCR mix included: 15 µL X2 Dream Taq Master Mix (Inqaba Biotechnological Industries Ltd, Pretoria, South Africa), the forward and reverse primers at a concentration of 0.4 µM each and 50 ng of the extracted DNA template. The PCR conditions were as follows: Initial denaturation at 95°C for 5min; denaturation at 95°C for 30 s; annealing at 50°C for 40 s; extension at 72°C for 50 s for 35 cycles then final extension at 72°C for 5 min. The products were determined by electrophoresis on 1% agarose gel at 120V/25 min and viewed on an ultra violet trans-illuminator (PREP ONETM Sapphire, EmbiTec) for a 120 bp product size.
Amplification of seb genes for S. aureus strains
The seb genes from the suspected enterotoxigenic S. aureus isolates were amplified using the seb primers (Table 1) on a DNA thermal cycler (GeneAmp PCR system) ABI 9700 Applied Biosystems, SA) at a final volume of 40 µl for 35 cycles. The PCR mix included: 20 X2 Dream Taq Master Mix supplied by Inqaba Biotechnological Industries Ltd Pretoria, SA, the primers at a concentration of 0.4 µM each and 50 ng of the extracted DNA template. The PCR conditions were as follows: Initial denaturation at 95°C for 5 min; denaturation at 95°C for 30 s; annealing at 48°C for 30 s; extension, 72°C for 30 s for 35 cycles and final extension, 72°C for 5 min. The product was separated by electrophoresis on a 1% agarose gel at 120V/25 min and visualized on a UV trans-illuminator (PREP ONETM Sapphire, EmbiTec) for a 409 bp product size.
Antimicrobial susceptibility tests for isolated strains of S. aureus in ready-to-eat edible larvae and land snail meats
A Kirby-Bauer method as described by Cheesbrough (2010) was employed to test the antibiotic susceptibility pattern of confirmed Staphylococcus spp. Eleven commercially used antibiotics (Rapid labs, UK and Oxoid, Bastingstoke, UK) were employed; namely: amoxycillin/clavulanate or augmentin (AUG) 30 µg, cefoxitin (FOX) 30 µg, ceftazidime (CAZ) 30 µg, ceftriaxone (CTR) 30 µg, cefuroxime (CRX) 30 µg, cloxacillin (CXC) 5 µg, gentamicin (GEN) 10 µg, erythromycin (ERY) 5 µg, ofloxacin (OFL) 5 µg, oxacillin (OX) 1 µg and vancomycin (VA) 30 µg. The tests were performed by standard disk diffusion technique on Mueller Hinton agar (TM, Rajasthan, India) with the turbidity standard (0.5 McFarland). The plates were incubated aerobically at 35°C for 16 to18 h. After incubation, test plates were examined for confluent growth and the diameter of each zone of inhibition was measured in millimetre (mm). Each inhibition zones diameter was interpreted using interpretive charts from WHO (CLSI, 2015).
Statistical analysis
The data obtained were analysed using descriptive statistics; one-way ANOVA and post Hoc Tukey test for significance P< 0.05. Proximate values of both food samples were in triplicates and analysed using Student T Test (p<0.05) to show their statistical significances. Some data were expressed in percentages, interpreted with tables using Microsoft Excel 2007.
Proximate values of ready-to-eat edible larvae and land snail meats
The mean proximate compositions of both ready-to-eat foods showed great differences between the nutritive values of edible larvae and snail meats (Table 2). The values of crude protein (26.12%), crude lipid (18.90%), fibre (5.12%), ash (3.40%) and total carbohydrate (32.99%) for edible larvae were higher compared to crude protein content (13.61%), lipid content (3.88%), fibre (2.01%), ash (1.11%) and total carbohydrate (19.79%) for snail meats with the exception of moisture content of snail meats (59.60%) which was much higher than edible larvae moisture content (13.47%).
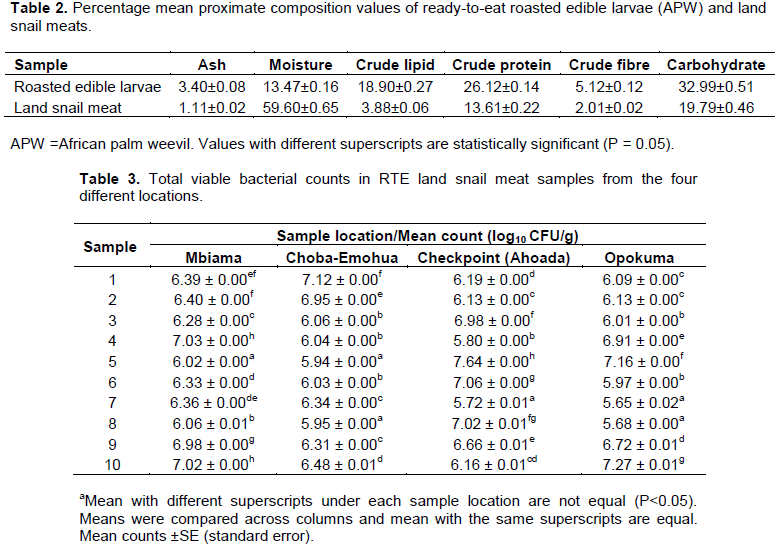
Total viable bacterial counts of ready-to-eat land snail meats and edible larvae
The mean total viable bacterial count (TVC) of RTE snail meats is shown in Table 3. The result showed that TVC (log10 cfu/g) in Ahoada was the highest (5.72±0.01 to 7.64±0.0), followed by Opokuma (5.65±0.02 to 7.27±0.1), Choba-Emohua (5.94±0.01 to 7.12±0.0) and the lowest was Mbiama (6.02±0.0 to 7.03±0.0). Table 4 shows the mean TVC of the RTE edible larvae. The TVC (log CFU/g) showed that Opokuma had the highest count (6.72±0.01 to 9.01±0.01), followed by Tombia (6.29±0.0 to 8.43±0.0), Mbiama (5.90±0.03 to 8.26±0.0) while Ekeki had the lowest count (5.60±0.05 to7.47±0.0).
Total staphylococcal counts of the ready-to-eat land snail meats and edible larvae
The mean staphylococcal count ofRTE snail meats for Mbiama, Choba-Emohua, Ahoada and Opokuma is shown in Table 5. Staphylococcal counts in log CFU/g were the highest in Opokuma (3.00±0.57 to 6.54±0.0) and the lowest were Choba-Emohua and Ahoada (3.60±0.05 to 5.91±0.01) and (3.00±0.57 to 5.91±0.0), respectively. Means within column with the same superscript are not significantly different (p=0.05). The mean total staphylococcal counts for edible larvae were the highest in Opokuma (4.51±0.0 to 8.62±0.01) log CFU/g and Ekeki had the lowest counts (3.70±0.1 to 6.76±0.0) log CFU/g, while Tombia and Mbiama staphylococcal counts (log10 CFU/g) ranged from (3.00±0.05 to7.45±0.0) and (3.28±0.0 to7.08±0.0), respectively (Table 6).
Interpretation of sanitary quality of the ready-to-eat foods edible larvae and land snail meats
In Table 7, the sanitary quality of the ready-to-eat food samples were interpreted with consideration to microbiological guidelines for ready-to-eat foods; category 5 (cooked foods chilled but with some handling prior to sale or consumption). All 80 analysed samples of both RTE edible larvae and snail meats revealed that 58.75% (47 of 80) of both samples were below the acceptable limit (105 to<107) CFU/g while 41.25% (33 of 80) of the samples were unsatisfactory (≥107 CFU/g). Total staphylococcal counts of the samples showed that 7. 5% (6 of the 80) were within acceptable limit (< 20 CFU/g), 27. 50% (22 of 80) at borderline limit (20 to ≤104) CFU/g and 65.00% (52 of 80) were unsatisfactory (>104 CFU/g). The RTE edible larvae from Opokuma and Tombia were the most unsatisfactory for consumption whereas the RTE snail meats samples were mostly at the borderline (105 to<107 CFU/g) (Health Protection Agency, 2009).


Detection of Staphylococcus spp. in the RTE roasted edible larvae and land snail meats
Out of the total of 80 samples (40 each) of the RTE edible larvae and snail meats analyzed, 74 (92.5%) and 47 (58.75%) of both RTE roasted edible larvae and land snail meats were positive for Staphylococcus spp. and S. aureus, respectively.
Molecular characterization of Staphylococcus spp. isolated from both ready-to-eat foods
Detection of enterotoxins A and B genes in the isolated Staphylococcus using PCR
Gel electrophoresis of seb genes from the isolated Staphylococcus strains: The agarose gel electrophoresis of amplified seb genes obtained when the DNA of S. aureus isolates were subjected to PCR using seb gene primers is as shown in Figure 3. A product size observed was 409 bp for seb genes. Lane L represents the molecular ladder lanes; N5, N7, and N15 showed positive on the seb gene bands.
Gel electrophoresis of sea genes from isolated Staphylococcus strains: The agarose gel electrophoresis of amplified sea genes obtained when the DNA of Staphylococcus isolates were subjected to PCR using sea gene primers is as shown in Figure 4. A product size observed was 120 bp for sea genes. Lane L represents the 100 bp molecular ladder lanes; N3, N5, N6, N7, N12, N14 and N15 showed positive on the sea gene bands.
Distribution of staphylococcal enterotoxins sea and seb genes in both ready-to-eat foods: In Table 8, Staphylococcus strains in both ready-to-eat foods produced sea and/or seb. SISA and OPSMI codes for S. aureus O326 and S. warneri JRT4, respectively harboured only sea virulent gene each. Also, 3/8 of S. aureus strains (with isolate codes ES3MS2a, SIMS3a and TS2Ma1) isolated from RTE edible larvae harboured both sea and seb virulent genes and isolated from Mbiama, Ekeki and Tombia, respectively. Other 2 strains (represented as S1MS2 and TS2MS1a) isolated from edible larvae produced only sea virulent genes and were obtained from Ekeki and Tombia, respectively. No seb virulent gene occurred singly in the 13 S. aureus strains.
Antibiotic resistance pattern of tested Staphylococcus spp. in both ready-to-eat foods: The S. aureus strains from both RTE edible larvae and snail meats were tested with 11 antibiotics. Gentamicin, ofloxacin and erythromycin were most active against all S. aureus strains at 100%. The resistance to ceftazidime, augmentin (amoxycillin-clavulanate) and cloxacillin was highest (100%). They showed varying resistance against ceftriaxone (57.14%), cefuroxime (28.57%), vancomycin (42.86%), oxacillin (42.86%) and cefoxitin (42.86%) (Figure 5).
Multiple-antibiotic resistance profile of Staphylococcus spp. isolated from the RTE samples: Staphylococcus spp. showed varying multiple resistances to antibiotics as presented in Table 9. Staphylococcus warneri JRT4 obtained from Opokuma had the highest percentage resistance of 63.6% to multiple antibiotics, followed by S. aureus O326 obtained from Choba-Emohua with 54.5%. S. aureus O326 and S. aureus P3-1 isolated from Mbiama and Ekeki, respectively showed the least multiple resistances of 27.3%.
In this study, proximate composition, microbiological qualities of the ready-to-eat edible larvae (R. phoenicis) and land snail (A. marginata) meats, presence of virulent genes and antibiotic sensitivities of S. aureus were investigated.
The proximate compositions of the ready-to-eat foods indicated that both RTE foods are sufficient in nutrients and edible larvae have more nutritional values compared to land snail meats. A number of authors have reported that edible larvae are very excellent sources of protein (Banjo et al., 2006; Opara et al., 2012; Amadi et al., 2014; Ebenebe and Okpoko, 2015) in comparison with proteins in chicken (24.96%), pork (16.57%), goat meat (20.14%) and beef (18.81%) (Afolabi et al., 2017). The findings of this study revealed that the protein content of RTE snail meats (13.6%) was less than the protein contents of 3 diverse kinds of unprocessed land snail meats ranging from 15.44 to 72.64% (Adegoke et al., 2010). The proximate value of lipid in this study is comparable to19.54% reported for the same kind of larvae by Okaraonye and Ikewuchi (2008). The lipid content of pork (36.87%) showed great disparity to lipid contents of edible larvae and snail meats (Afolabi et al., 2017). The lipid value (3.88%) for RTE snail meats in the current study is more than the crude lipid contents range (1.15 to 1.52 %) reported by Adegoke et al., (2010). Akinnusi (2002) stated that the low content of fat and low cholesterol level make snail meat a remedy for vascular diseases such as heart attack, cardiac arrest, hypertension, stroke, high blood pressure and other fat related ailments. Contrary to reports by Opara et al., (2012) that edible larvae are a poor energy giving foods, the carbohydrate content of edible larvae in this study was high (32.99%). The moisture content of these foods evaluated reveals their stability and susceptibility to contamination by organisms (Uraih and Izuagbe, 1990). The low fibre content of RTE snail meats implies that they can serve as roughage. Edible larvae are adequate source of diet roughage for bowel peristalsis (Okaraonye and Ikewuchi, 2008). The nutritive values of both foods show they are considerate reserves of protein, lipid, carbohydrate and moisture.
Most of the food samples had TVC exceeding 105 CFU/g as has been reported by other authors in Nigeria, Congo and Sri Lanka (Ekrakene and Igeleke, 2007; Ebenebe and Okpoko, 2015; Makelele et al., 2015; Wimalasekara, 2016). The TVC was the highest in RTE snail meats from Ahoada (5.72 to 7.64 log10 CFU/g), while for RTE edible larvae, Opokuma had the highest counts (6.72 to 9.01 log10 CFU/g). Total presumptive staphylococcal counts in snail meats and edible larvae were both highest in Opokuma having counts ranging from 3.0 to 6.54 log10 CFU/g and 4.51 to 8.62 log10 CFU/g, respectively. In line with the TVC for RTE snail meats, Nyoagbe et al. (2016) reported TVC in land snail species ranging from 6.00 to 8.00 log10 CFU/g while the TSC range from 2.00 to 7.00 log10 CFU/g. The enormous degree of contamination in these ready-to-eat foods may not be unrelated to the processing environment, initial microbial loads, extent of exposure at selling points, and food handling by food vendors. Staphylococci counts in cooked foods suggest improper and repeated food handling after food processing. High counts of Staphylococcus spp. in cooked foods renders them unfit for man consumption.
The results of virulence gene detection revealed an 8.64% (7 of 81) harbours sea genes, 6 S. aureus and S. warneri strains while 3.70% (3 of 81) harbours the sea genes, 3 S. aureus strains. This finding is not comparable to the research findings carried out in Gorgan city of Iran (Kamarehei et al., 2013) which reported that out of 170 S. aureus isolates, 60.6% contained sea genes while 27.1% carried seb genes, but followed the same trend with the sea genes dominating. Leke et al. (2017) also reported that 24.1% S. aureus isolated from breast milk, had sea gene. The sea gene is the most common toxin associated with staphylococcal food poisoning and seb is also linked to food poisoning as well as an inhaled bio-weapon (Balaban and Rasooly, 2000; Pinchuk et al., 2010). The study further revealed that the S. aureus produced either sea and/or seb genes. S. aureus P3-1 produced only sea genes whereas only S. aureus O326 obtained from both RTE foods produced sea and/or seb genes. S. aureus O326 was the most distributed strain in the foods across the sampled locations. S. aureus O326 and P3-1 present in the edible larvae at Ekeki, while S. warneri JRT4 in Opokuma. The virulent genes presence demonstrated the potential toxigenicity and pathogencity of the Staphylococcus isolates. The sea and seb are classical types of SEs and influence emetic activity. The sea genes were isolated both in edible larvae and snail meats while seb-carrying strains were only common in edible larvae. This occurrence is possibly caused by epidemiological factors like type of strains and environment. Delayed processing, inadequate refrigerating, poor personal cleanliness and post-process contamination are related to the growth of S. aureus strains harbouring SE and ET genes (Leke et al., 2017).
Previous reports confirmed the enterotoxigenicity of other Staphylococcus spp. other than S. aureus (Jay, 1992). In this study, S. warneri JRT4 harboured staphylococcal enterotoxins sea genes and biochemically was positive to coagulase. According to Becker et al., (2001), few coagulase positive Staphylococcus intermedius strains harbour SEs present in food.
Antimicrobial resistance from foods and water sources are of global concern (Kumar et al., 2005). There were varying degrees of resistance of the Staphylococcus strains to the commercial antibiotics. The virulent genes bearing Staphylococcus were 100% resistant to augmentin, ceftazidime and cloxacillin but showed varying resistance against ceftriaxone (57.14%), cefuroxime (28.57%), vancomycin (42.86%), oxacillin (42.86%) and cefoxitin (42.86%). Overall, they were 100% sensitive to gentamicin, ofloxacin and erythromycin. High degree of sensitivity of different organisms to ofloxacin and gentamicin has been previously reported (Okonko et al., 2008; Mordi and Momoh, 2009; Umofia, 2012; Afolabi et al., 2017). Agbo et al. (2016) reported that S. aureus isolated from street foods in Calabar had varying resistance to ciprofloxacin (20%), gentamicin (10%), and levofloxacin (5%). Kumar et al. (2009) reported higher resistance of S. aureus against erythromycin (87.5%), while Achi and Madubuike (2007) reported lesser resistance against erythromycin (0.83%) which is comparable to the findings of this study. According to Harakeh et al. (2005) and Guven et al., (2010), meals prepared in the street provide a suitable culture medium for S. aureus strains impervious to various antibiotics and transmitted to people via street foods that are contaminated. S. aureus are usually regarded to be resistant to antibiotic therapy, as a result of exopolysaccharide obstruction and their sites inside micro abscesses which reduce the actions of drugs (Gundocan et al., 2006). Food handlers and the surrounding that harbour antibiotic resistant S. aureus strains transfer their resistant genes to the sustenance likely because of their poor cleanliness and sanitation or the misuse of antibiotic administrations. S. aureus O326 and S. warneri JRT4 isolated from RTE snail meat from Choba and Opokuma, respectively showed highest multiple resistances to the selected antibiotics confirming earlier report by Waters et al. (2011) that multidrug resistance S. aureus occurs often.
In this finding, ofloxacin, a second era quinolones, erythromycin and gentamicin, an aminoglycosides are the most dynamic broad spectrum antibacterial agents to combat ailments brought about by the Staphylococcus.
The findings of this investigation demonstrated that RTE foods examined are excellent sources of essential nutrients, with edible larvae having more nutritional benefits compared to land snail meats. The study showed that 41.25% samples tested had total viable above the acceptable microbiological limit (≥107 CFU/g) for ready-to-eat vended foods. Staphylococcus spp. carried the sea and/or seb virulence genes affirming their ability to initiate disease conditions for commuters who patronize these vendors. The strains of S. aureus and S. warneri isolated from the RTE edible larvae and snail meats showed various dimensions of resistance pattern to the commercial antibiotics utilized in global reports, hence the requirement for a superior and realistic approach to ensure road side food vendors comply with standard food safety practices.
The authors have not declared any conflict of interests.
The authors are grateful to the technical staff of Emadavistic Medical and Research Laboratory, Osaks House, East-West Road Port Harcourt and Department of Medical Laboratory Science, Niger Delta University, Nigeria for their assistance.
REFERENCES
|
Achi OK, Madubuike CN (2007). Prevalence and Antimicrobial resistance of Staphylococcus aureus isolated from Retail Ready-to-eat Foods in Nigeria. Research Journal of Microbiology 2(6):516-523.
Crossref
|
|
|
|
Adegoke AA, Komolafe AO (2009). Multi-drug resistant Staphylococcus aureus in clinical cases in Ile-Ife, South-west Nigeria. International Journal of Medical Sciences 1(3):068-072.
|
|
|
|
|
Adegoke AA, Adebayo-Tayo CB, Inyan UC, Aiyegoro AO, Komolafe OA (2010). Snail as meat source: Epidemiological and Nutritional Perspective, Journal of Microbiology and Antimicrobial 2(1):001-005.
|
|
|
|
|
Afolabi FT, Arowesebe AR, Adeyemi SM (2017). Antibiotic Susceptibility pattern and Plasmid Profiling of some isolates obtained from raw meats sold in Ibadan, Metropolis. Journal of Microbiology and Biotechnology Research 7(4):13-22.
Crossref
|
|
|
|
|
Agbo BE, Udoekong NS, Ozumba RE (2016). Incidence of Staphylococcus aureus in Street vended food sold in Calabar Municipality, Nigeria. Journal of Biopesticides and Agriculture 3(1):31-40.
|
|
|
|
|
Agbonlahor DE, Imoyera PI, Igumboe EO, Akhabue EE, Ekundayo PA, Orhue DT, Obasuyi BO, Osuide OO (1994). The bacteriology of edible giant Africa land snail (Archachatina marginata) commonly found in Southern part of Nigeria. Journal of Medical Laboratory Science 4:26-32.
|
|
|
|
|
Ajayi SS, Tewe SO, Milligan JK (2009). Influence of seasonality on aestivation and behavior of the forest giant land snail, Archachatina marginata (Swaison). Bulletin of Animal Health Protection 28:328.
|
|
|
|
|
Akinnusi O (2002). Introduction to snails and snail farming. Triolas Publishing Company, Abeokuta P 70.
|
|
|
|
|
Amadi EN, Kiin-Kabari DB, Kpormon LB, Robinson VKK (2014). Microbial flora and Nutritional Composition of Adult Palm wine Beetle (Rhynchophorus phoenicis). International Journal of Current Microbiology and Applied Science 3(11):189-192.
|
|
|
|
|
Association of Official Analytical Chemists (AOAC) (2000). Official methods of Analysis of AOAC International, 17th Edition, II. Washington, DC, USA.
|
|
|
|
|
Arene FO, Ibanga ES, Asor JE (1999). Freshwater snail and Crab intermediate hosts of Paragonimus species in two rural communities in Cross River Basin, Nigeria. Global Journal of Pure and Applied Science 5:184-187.
|
|
|
|
|
Balaban N, Rasooly A (2000). Staphylococcal enterotoxins. International Journal of Food Microbiology 61(1):1-10.
Crossref
|
|
|
|
|
Banjo AD, Lawal OA, Songonuga EA (2006). The nutritional value of fourteen species of edible insects in South-western Nigeria. African Journal of Biotechnology 5(3):298-301.
|
|
|
|
|
Becker K, Keller B, Von EC, Bruck M, Lubritz G, Etiene J, Peters G (2001). Enterotoxigenic potential Staphylococcus intermedius. Applied Environmental Microbiology 67(12):5551-5557.
Crossref
|
|
|
|
|
Chang SK (2003). Protein analysis in Food. 3rd Edition. Kluwe Academic Plenum Publishers: New York, USA pp. 134-135.
|
|
|
|
|
Cheesbrough M (2010). District Laboratory Practice in Tropical Countries, Part 1. Cambridge University Press.
|
|
|
|
|
CLSI (2015). Performance Standards for Antimicrobial Disk Susceptibility Tests; Approved Standards-12th Edition. Clinical and Laboratory Standards Institute. Wayne, USA.
|
|
|
|
|
Daniel EO, Onulide AA (2017). Isolation of Multidrug resistant Listeria monocytogenes from processed Rhynchophorus phoenicis Larvae. Bioresearch Communications 3(1):340-344.
|
|
|
|
|
Ebenebe CI, Okpoko VO (2015). Microbiological Quality of Raw and Roasted African Palm Weevil (Rhynchophorus phoenicis) consumed in South-Eastern Nigeria. Animal Research International 12(2):2159-2165.
|
|
|
|
|
Ekrakene T, Igeleke CL (2007). Microbial isolates from the Roasted Larva of the Palm Weevil Rhynchophorus phoenicis [F] from Edo and Delta States of Nigeria. Australian Journal of Basic and Applied Sciences 1:763-768.
|
|
|
|
|
Eruteya OC, Odunfa SA, Lahor J (2014). Listeria spp. in Raw Cow and Goat Meat in Port Harcourt, Nigeria. British Biotechnology Journal 4(2):204-214.
Crossref
|
|
|
|
|
Fagbuaro O, Oso JA, Edward JB, Ogunleye RF (2006). Nutritional status of four species of giant land snails in Nigeria. Journal of Zhejiang University Science B 7:686-689.
Crossref
|
|
|
|
|
Fogoh JM, Aurele AA, Philippe L, Patrice L (2015). Exploitation, Trade and Farming of Palm weevil grubs in Cameroon. Working paper 178, Bogor, Indonesia; CIFOR.
|
|
|
|
|
Gundocan N, Citak S, Turan E (2006). Slime production, DNase activity and antibiotic resistance to Staphylococcus aureus isolated from raw milk, pasteurized milk and ice cream samples. Food Control 17(5):389-392.
Crossref
|
|
|
|
|
Guven K, Mutlu MB, Gulbandilar A, Cakir P (2010). Occurrence and characterization of Staphylococcus aureus isolated from meat and dairy product consumed in Turkey. Journal of Food Safety 30(1):196-212.
Crossref
|
|
|
|
|
Harakeh S, Yassine H, Gharios M, Barbour E, Hajjar S, El-Fadel M, Toufeili I, Tannous R (2005). Isolation, molecular characterization and antimicrobial resistance of Salmonella and Escherichia coli isolates from meat-based fast food in Lebanon. Science of the Total Environment 341(1-3):33-44.
Crossref
|
|
|
|
|
Harrigan WF, McCance ME (1990). Laboratory Methods in Food and Dairy Microbiology (8th Edition). Academic Press Inc.: London pp. 7-23; 286-303.
|
|
|
|
|
Health Protection Agency (2009). Guidelines for Assessing the Microbiological Safety of Ready-to-eat Foods. London: Health Protection Agency.
|
|
|
|
|
Herbert DG, Kilburn RN (2004). Field guide to the land snails and slugs of eastern South Africa. Natal Museum, Pietermaritzburg.
|
|
|
|
|
James CS (1995). Analytical Chemistry of Foods. Chapman and Hall: New York, USA pp. 41-75.
Crossref
|
|
|
|
|
Jay JM (1992). Staphylococcal gastroeneteritis. In: Modern Food Microbiology (Nostrand, V. editions). 4th Edition. New York, NY, USA pp. 455-478.
Crossref
|
|
|
|
|
Kamarehei F, Ghaemi EA, Dadgar T (2013). Prevalence of enterotoxin a and b genes in Staphylococcus aureus isolated from clinical samples and healthy carriers in Gorgan City, North of Iran. Indian Journal of Pathological Microbiology 56(3):265-268.
Crossref
|
|
|
|
|
Kumar HS, Parvathi A, Karunasagar I (2005). Prevalence and antibiotic resistance of Escherichia coli in Tropical sea food. World Journal of Microbiology and Biotechnology 21:619-623.
Crossref
|
|
|
|
|
Kumar JD, Negi YK, Gaur A, Khanna D (2009). Detection of virulence genes in Staphylococcal aureus isolated from paper currency. International Journal of Infectious Diseases 13(6):450-455.
Crossref
|
|
|
|
|
Lancette GA, Bennett RW (2001). Staphylococcus aureus and staphylococcal enterotoxins. In: Downes FP, Ito K (edition). Compendium of Methods for the Microbiological Examination of Foods. 4th Edition. Washington, American Public Health Association.
Crossref
|
|
|
|
|
Le Loir Y, Baron F, Gautier M (2003). Staphylococcus aureus and food poisoning. Genetics and Molecular Research 2(1):7-28.
|
|
|
|
|
Leke A, Goudjil S, Mullie C, Grognet S, Biendo M (2017). PCR detection of Staphylococcal enterotoxins genes and exfoliative toxin genes in methicillin resistant and Methicillin susceptible Staphylococcus aureus strains from raw human breast milk. Clinical Nutrition Experimental 14:26-35.
Crossref
|
|
|
|
|
Makelele LK, Kazadi ZA, Oleko RW, Mpalang NK, Ngbolua NK, Gedeon BN (2015). Microbiological quality of food sold by street vendors in Kisangani, Democratic Republic of Congo. African Journal of Food Science 9(5):285-290.
Crossref
|
|
|
|
|
Mordi R, Momoh M (2009). Incidence of Proteus species in wound infections and their sensitivity pattern in the University of Benin Teaching Hospital. African Journal of Biotechnology 8(5):725-730.
|
|
|
|
|
Nwuzo AA, Iroha IR, Moses IB, Ugbo EN, Agumah NB, Orji J, Okonkwo EC, Oke B, Ogene L (2016). Isolation and Characterization of Bacterial Species associated with Edible Snails (Achatina achatina) Sold in Major Markets within Abakaliki Metropolis. Biolife 4(3):494-497.
|
|
|
|
|
Nyoagbe LM, Appiah V, Nketsia TJ, Larbi D, Adeji I (2016). Evaluation of African giant snails (Achatina and Archachatina) obtained from markets (wild) and breeding farms. African Journal of Food Science 10(7):94-104.
Crossref
|
|
|
|
|
Okaraonye CC, Ikewuchi JC (2008). Rhynchophorus phoenicis (F) Larva Meal: Nutritional value and health implications. Journal of Biological Sciences 8(7):1221-1225.
Crossref
|
|
|
|
|
Okonko IO, Ogunjobi AA, Fajobi EA, Onga BA, Babalola ET, Adedeji AO (2008). Comparative studies and different assessment of ready-to-eat frozen sea foods processed in Ijita, Olapa Lagos State, Nigeria. Journal of African Biotechnology 7(16):2898-2901.
|
|
|
|
|
Opara MN, Sanyigha FT, Ogbuewu IP, Okoli IC (2012). Studies on the production trend and quality characteristics of palm grubs in the Tropical rainforest zone of Nigeria. Journal of Agricultural Technology 8(3):851-860.
|
|
|
|
|
Oranusi US, Braide W (2012) A Study of Microbial Safety of Ready-To-Eat Foods vended on Highways: Onitsha-Owerri, South-Eastern Nigeria. International Research Journal of Microbiology 3(2):66-71.
|
|
|
|
|
Pinchuk IV, Beswick EJ, Reyes VE (2010). Staphylococcal enterotoxins. Toxins 2(8):2177-2197.
Crossref
|
|
|
|
|
Tambe TJ, Riolo P, Okolle NJ, Isidoro N, Fanciulli PP, Dallai R (2013). Sexual size differences and color polymorphism of Rhynchophorus phoenicis in the Southwest region of Cameroon. Bulletin of Insectology 66(1):153-159.
|
|
|
|
|
Thomas D, Chou S, Dauwalder O, Lina G (2007). Diversity of Staphylococcus aureus enterotoxins. Chemical Immunology and Allergy 93((1):24-41.
Crossref
|
|
|
|
|
Umofia I (2012). Antibiotic Susceptibility studies of some bacterial isolates from packaged milk of marketed in Zaria, Nigeria. M.Sc Thesis. Department of Pharmaceutics and Pharmaceutical Microbiology, Ahmadu Bello University, Zaria, Nigeria.
|
|
|
|
|
Uraih N, Izuagbe Y (1990). Public Health, Food and Industrial Microbiology. 1st Edition. Uniben Press, Nigeria 54 p.
|
|
|
|
|
USDA (2006). National Nutrient Standard Reference, Release 19. Available from
View. (Accessed February 5, 2018).
|
|
|
|
|
Vendruscolo JW, Waldrich TL, Saikawa GIA, Pelayo JS, Kbayashi RKT, Nakazato G, Rocha SPD (2017). Novel Multiplex PCR for detection of diarrheagenic Escherichia coli strains isolated from stool and water samples. Genetics and Molecular Research 16(3):gmr16039760.
Crossref
|
|
|
|
|
Wachukwu CK, Thomas CN, Kigigha LT (2002). Microorganisms associated with Palm weevil larvae (Ryhnchophorus phoenicis). Journal of Dairying Food and Home Science 21(1):44-47.
|
|
|
|
|
Waters AE, Contente-Cuomo T, Buchhagen J, Liu CM, Watson L, Pearce K, Foster JT, Bowers J, Driebe EM, Engelthaler DM, Keim PS, Price LB (2011). Multidrug-resistant Staphylococcus aureus in US meat and poultry. Clinical Infectious Diseases 52(15):1227-1230.
Crossref
|
|
|
|
|
Wieneke AA, Roberts D, Gilbert RJ (1993). Staphylococcal food poisoning in the United Kingdom, 1969-90. Epidemiological Infections 110(3):519-531.
Crossref
|
|
|
|
|
Wimalasekara G (2016). Microbiological quality of ready-to-eat meat-based foods available in temporary food outlets in Gall Face Green, Colombo, Sri Lanka. International Journal of Agriculture, Forestry and Plantation 4:38-44.
|
|
|
|
|
Womeni HM, Tiencheu B, Linder M, Nabayo EMC, Tenyang N, Mbiapo FT, Villeneuve P, Fanni J, Parmentier M (2012). Nutritional value and effect of cooking, drying and storage process on some functional properties of Rhynchophorus phoenicis. International Journal of Life Science and Pharma Research 2(3):203-219.
|
|