Full Length Research Paper

ABSTRACT
ARTAVOL® is an herbal product derived from de-artemisinised extracts of Artemisia annua combined with lemon grass extract and avocado seed powder as a base locally used in Uganda for malaria prevention. This study assessed the immunomodulatory activity of the aqueous extract of Artavol® in Plasmodium berghei infected mice. The immunomodulatory activity of the aqueous extract of Artavol® was evaluated on Chloroquine-sensitive P. berghei ANKA-infected mice using 7 days test at 15, 30, and 60 bmg/kg with Pyrimethamine (1.25 mg/kg) as positive control; and distilled water as negative controls. Seven days post – infection, mice were euthanized and the spleen collected from each mouse under sterile conditions. Phenotypic and functional characterization was T-cell surface and intracellular markers was done by CytofleX LX flow Cytometer, and the data obtained were analyzed using FlowJo version 10.7.1 software. All the extracts (15-60 mg/kg) doses showed a significant increase in the frequency of monocytes (p=0.0078, p=0.0084, and p<0.0001, respectively) compared to the negative control. The extract at 30 and 60 mg/kg showed a gradual significant increase (p<0.0001 and p< 0.0001, respectively) in the frequency of natural killer cells (NK cells) compared to the negative control. The aqueous extract of ARTAVOL® possesses variable immunomodulatory effects and showed ability to increase the frequency of monocytes and natural killer cells. IFN-γ and TNF-α levels showed a significant increase suggesting the extract induced their responses.
Key words: Artavol®, Immunomodulatory effect, Plasmodium berghei.
INTRODUCTION
The world health organization promotes the use of chemoprophylactic drugs to treat and prevent malaria in endemic areas (World Health Organization, 2015). Chemoprophylaxis has been effective in preventing morbidity and mortality from malaria among high-risk groups such as pregnant women, infants and children (Tizifa et al., 2018) hence, tourists travelling to endemic and high-risk areas are advised to take prophylactic antimalarial agents (Schlagenhauf et al., 2019; Shellvarajah et al., 2017). World health organization has also recommended the use of recombinant protein-based malaria vaccine (RTS, S/AS01) among children in sub-Sharan Africa and other regions with moderate to high Plasmodium falciparum transmission (Trottier and Elliott, 2021). Currently, all available chemoprophylactic anti-malarial agents do not provide 100% protection and using these agents for long period might interfere with the development of naturally acquired immunity and facilitate the emergence and spread drug resistant strains (Greenwood, 2010). With the problems of increasing levels of drug resistance among conventional antimalarials, herbal medicines could be an important and sustainable source of treatment of malaria parasites (Willcox and Bodeker, 2004).
Medicinal plants andl products have been used for prophylaxis and treatment of malaria in sub-Saharan Africa for years (Chinsembu, 2015). Use of herbal remedies has also been embraced in many developed and developing countries as complementary and alternative medicines (Ekor, 2014).
In Uganda, a number of medicinal plants used to treat malaria have been reported (Ajayi et al., 2020; Tugume et al., 2016). ARTAVOL® is one of the prophylactic antimalarial herbal products used locally in Uganda (Mesa et al., 2017; Ogwang et al., 2011). It is composed of Avocado seed extracts, Lemon grass, and Artemisia annua without artemisinin.
The ARTAVOL® powder can be mixed in hot water, porridge or hot milk and taken as a beverage to prevent malaria (Ogwang et al., 2011). Although ARTAVOL® has been proven to exhibit antimalarial activity, there is need to assess its immunomodulatory effects. This study, therefore, assessed the immunomodulatory responses of the aqueous extract of ARTAVOL® in Swiss albino mice infected with P. berghei ANKA parasites.
MATERIALS AND METHODS
Study design
The study employed a controlled in vivo experimental design in which the effects of ARTAVOL® aqueous extract on the cell Mediated immune response in P. berghei infected mice was evaluated.
Malaria parasite strain
Chloroquine sensitive P. berghei ANKA MRA 311 was obtained from BEI resources, NIAID, NIH contributed by Thomas F. McCutchan. The vial containing 0.5 mL P. berghei ANKA strain parasite in dried ice was activated by thawing frozen cryovial in water bath at 35°C for 2 min. This was followed by wiping the outside of the vial with 70% ethanol before opening and 50 µL injected intraperitoneally into each mouse. The growth of the parasites was monitored by tail vein blood sampling and Giemsa-stained thin blood smear microscopically daily, starting on day -3 post infections.
Laboratory animals, care and ethical approval
Adult male and female Swiss albino mice between 20-30 g were obtained from the Animal Facility Research, Mbarara University of Science and Technology, Uganda. The mice were fed on grower pellets purchased from Kuyoyo Feeds, Uganda and had free access to water. The mice were acclimatized for two weeks before the experiments. The study was carried out between September 2021 to December 2021. All the animals in the experimental study were treated humanely according to the Organization for Economic Co-operation and Development (OECD) guidelines and American Psychological Association (Bedi and Krishan, 2020, Loue, 2020). ARTAVOL® product (Batch number: 005) was obtained from ARTAVOL Ltd (ARTAVAL Ltd. P.O. Box.34 Ntinda, Kampala- Uganda the manufacturer. Permission to use ARTAVOL® herbal product in the study was obtained from developers of the product. This study also received ethical clearance from Mbarara University Research and Ethics Committee with approval number 07/05-19 and the Ugandan National Council for Science and Technology registration number HS465ES.
Preparation of crude ARTAVOL extracts
ARTAVOL® extract was prepared using infusion method in which 60 mL of distilled water was boiled at 80°C, poured into 173 g powdered ARTAVOL®, and allowed to stand for 15 min. Thereafter, the extract was filtered, concentrated in vacuo at 55°C and freeze-dried. The aliquot of the stock solution of 5 mg/mL concentration was prepared and stored in a fridge below 4°C. The test doses of 15, 30, and 60 mg/kg was separately determined from the stock solution to have a final volume of 60, 120 and 240 µL and administered orally to already grouped mice.
Acute toxicity study of the crude ARTAVOL extracts
The median lethal dose (LD50) of ARTAVOL® was determined in vivo using the Lorke’s method (Lorke, 1983). In the first phase, nine mice were randomly divided into three groups of three mice each and each group received the extract at 10, 100 and 1000 mg/kg body weight orally. The mice were then observed for signs of adverse effects and mortality for the first 48 h and then for 12 more days. In the second phase following the same Lorke’s method, three mice were divided into 43 groups of 1 mouse each. The animals were given higher doses (1600, 2900, and 5000 mg/kg) of the test extract. The animals were then monitored for any sign of toxicity like stretching, rubbing of nose on the floor and wall of cage, change in body weight and mortality over a period of 24 hours and then 14 days. LD50 was calculated using the formula:
LD50 ? [Do × D100]1/2
Where Do = highest that caused no mortality and D100 = the lowest dose that produced mortality (Lorke, 1983).
Preparation of malaria parasite Inoculum
Chloroquine (CQ)-sensitive P. berghei ANKA parasites were activated in the mice according to BEI Resources user reference.. About 200 µL of the inoculum was injected into the donor mouse intraperitoneally and the parasites growth was monitored after 72 h by preparation of smear from the blood taken from the tail of infected mice. The parasites were maintained by continuous blood passage of the blood collected from the donor mouse into a new group of mice. A standard inoculum of 1×107 parasitized erythrocytes was prepared by dilution of blood collected through cardiac puncture from a donor mouse (> 30% parasitemia) with normal saline and administered intraperitoneally (200 µL) to each test mouse.
Determination of immune response
Thirty (30) mice were grouped into 5 groups of 6 mice each; Mice in Groups I – III were orally administered with ARTAVOL® (15, 30 and 60 mg/kg, respectively) by using oral cannula while the animals in Group IV were given Pyrimethamine at 1.25 mg/kg/day (positive control) and group V (negative control) received 200 µL/mouse/day distilled water for seven consecutive days (D1 – D7) respectively. On day 7, all the mice were inoculated with P. berghei parasites. On day 7 post infection, mice were euthanized and the spleen collected from each mouse under sterile conditions. The spleens were placed in RPMI 1640 (Thermo Fisher Scientific, Waltham, MA USA) media, crushed through a cell strainer. The cell suspension was washed twice using 1X PBS (washing buffer) and centrifuged at 1500 rpm for 10 min at 25°C. RBCs in the pellet were lysed using 2.5 mL of lysing buffer (Gibco, USA) for 2 min at room temperature. The lysing buffer was washed off and splenocytes counted using a hemocytometer machine. The splenocytes were cryopreserved in liquid nitrogen until use.
Splenocytes thawing and staining
Cryopreserved splenocytes were retrieved from liquid nitrogen at -196°C and immediately transferred to a preset 37°C water bath. Upon thawing, cells were washed in complete RPMI 1640 medium and kept overnight at 37°C in 5% carbon dioxide (CO2). Thereafter, the cell yield viability using trypan blue solution was determined. Cells were stained using 0.4% Trypan blue solution at a 1:1 dilution ratio. Samples with at least 70% viability were used for stimulation assays.
Cell surface staining
About 4.0 ×105 cells/mL were surface stained and incubated for 30 min with the following antibodies; CD3ε Brilliant Violet 510™ (Cat No. 100353), CD4 PE-Cy5(Cat No 100409), CD8b.2 APC/Fire™ 750(Cat. No. 140419), CD11b FITC (Cat. No. 101206), CD25 PE/Dazzle™ 594 (Cat No 102047), CD69 PE/Cyanine7 (Cat. No. 104512), CD279 (PD-1), Brilliant violet 421 (Cat. No 109121), and CD335 (NKp46) APC (Cat. No. 137608) in which 1 uL each of surface markers were added to the samples and incubated for 30 min at room temperature in the dark. The cells were washed twice by adding cell staining buffer and centrifuged at 1800 rpm for 5 min. The cells were acquired on a 19 color CytofleX LX flow Cytometer (Beckman coulter, New Jersey, USA). At least 100,000 events were recorded for analysis. Gating was standardized and set using fluorescence minus one control (FMOs).
Intracellular staining of cytokines
Following surface staining, the cells (splenocytes) were fixed by adding 0.5 mL of fixation buffer to each sample tube, incubated in the dark for 20 min at room temperature and then centrifuged at 1800rpm for 5 min and the supernatant discarded. The fixed cells were permeabilized by re-suspending in intracellular staining perm wash buffer (diluted to 1X in deionized water) and centrifuged at 1800 rpm for 5 min. A cocktail of intracellular cytokine antibodies was prepared by adding 1ul of each that is, IFN-γ BV605 (Cat. No 505839), TNF-αBV650 (Cat. No 506333), IL-2 BV785 (Cat. No. 503843), IL-4 PerCP Cy5.5 (Cat. No 504123), IL-10PE (Cat No. 505008), and IL-17A Alexa F700 (Cat. No 506914) in the tube. The antibody cocktail was added to each of the sample tubes and incubated for 20 min in the dark at room temperature. Cells were washed 2 times with 2 mL of Intracellular Staining Perm Wash Buffer and centrifuged at 1800 rpm for 5 min. The cells were then resuspended in 200 µL Cell Staining Buffer. The samples were acquired on the Cytoflex LX flow cytometer.
Compensations and fluorescent minus ones (FMOs) controls for each of the antibody fluorochromes were ran together with the optimization gains for every channel using unstained cells. Compensation calculations were done and applied to the samples. At least one hundred thousand events were acquired and recorded per sample. Data was exported for analysis using flowJo software.
Statistical analysis
Flow cytometry data were analyzed using FlowJo version 10.7.1 software (Becton Dickinson, New Jersey, USA). Statistical data were analyzed using Graph Pad Prism version 8.0.3 (Prism, 2018). Lymphocytes were carefully gated on a forward scatter and side scatter plot. The percentage of positive cells for each marker was determined on the basis of the numbers of viable lymphocytes. The frequencies of T cells expressing activation and memory markers were measured as a percentage of the CD4+ and CD8+ T cell parent cell populations. Bivariate dot plots or probability contour plots were generated upon data analysis to display the frequencies of and patterns by which individual cells co-express certain levels of cell surface antigen and intracellular cytokines. Results obtained were expressed as mean ± standard error of mean. Differences in the frequencies of the T cell subsets were evaluated using a One-way Analysis of Variance while the difference among the means was considered at 95% confidence interval using the post-hoc method of Dunnett's Multiple Comparison Test.
RESULTS
The acute oral toxicity test of aqueous ARTAVOL® extract caused no gross behavioral changes such as loss of appetite, paw licking, body temperature, calmness, locomotion, etc., and there was no mortality within 24 h as well as in the next 14 days, indicating that the LD50 values of the aqueous ARTAVOL® extract were greater than 5000 mg/kg in mice.
Gating strategy
A gating strategy was developed to analyze P. berghei activation marker expression on CD4+ and CD8 T-cells (Figure 1).
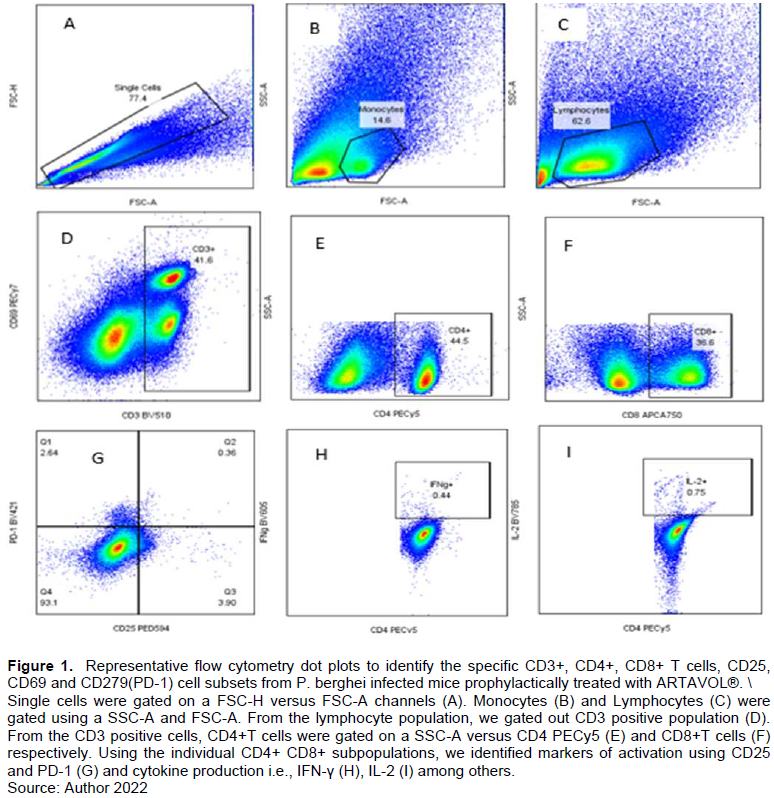
Effect of ARTAVOL® extract on CD4 T-cell response in P. berghei-Infected mice
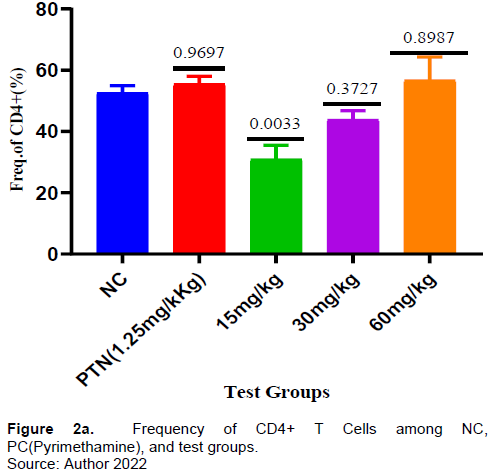
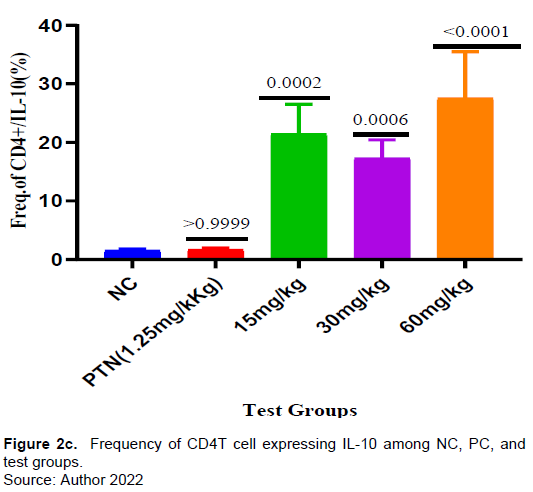
There was no increase in the frequency of CD4+ T-cells in all the test samples compared to both the positive and negative controls but the frequency of CD4+ T-cells increases as the test dose increases (Figure 2a). Interestingly, the percentage of CD4 T-cells expressing I L-10, was significantly higher in all the test groups compared to the negative control (Figure 2c).
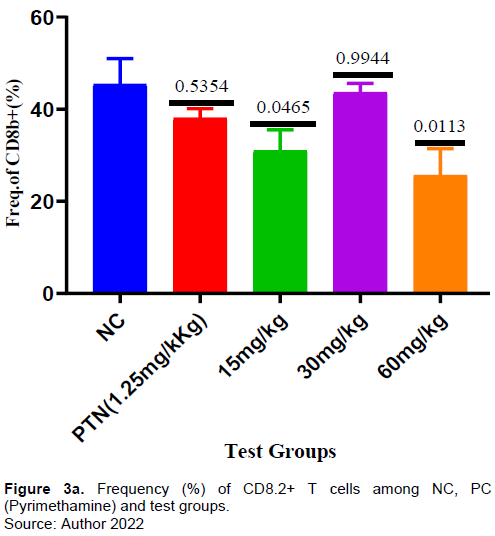
Effect of ARTAVOL® extract on CD8 T-cell response in P. berghei-Infected mice
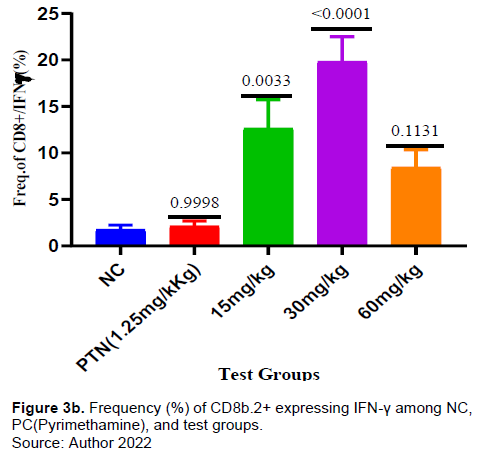
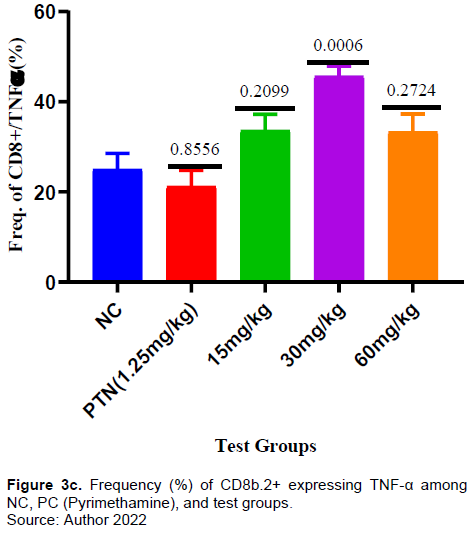
There was no significant increase in the frequency of CD8+ T-cells in the test compared to negative control (Figure 3a). However, the frequency of CD8 T-cells expressing IFN-γ for the test group 15mg and 30mg/kg was significantly higher compared to negative control (Figure 3b). The frequency of CD8+ T-cells expressing TNF-α for the test group 30 mg/kg was significantly higher (p = 0006) compared to negative control. There was increase in the frequency of CD8 T-cells expressing TNF-α in the other test doses (15 mg and 60 mg/kg) and positive control but it was not significant when compared with the negative control (Figure 3c). The frequency of PD1-expressing CD8+ T cells was higher in the test dose of 30 mg/kg, but lower in the test doses of 15 and 60mg/kg when compared to the negative control (Figure 3d).
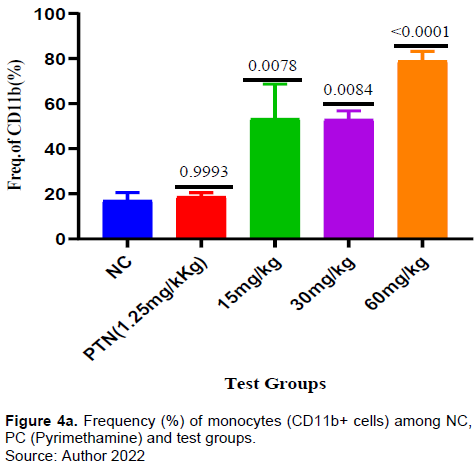
Effect of ARTAVOL® extract on CD11 (monocytes) response in P. berghei-Infected mice
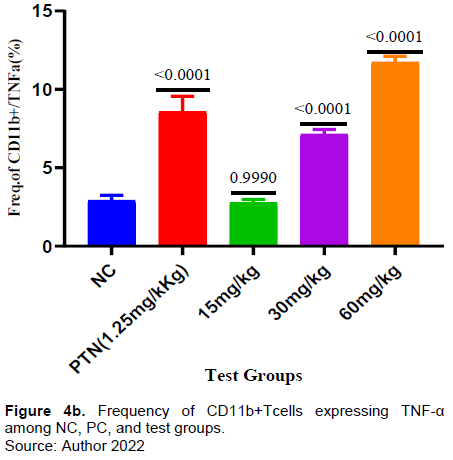
It was found that the frequency of CD11 cells (monocytes) was significantly higher in all the test groups when compared with the negative control. (Figure 4a). Similarly, there was a significant increase of monocytes expressing TNF-α in the test groups of 30mg and 60mg/kg and in the positive control when compared with the negative control. In test dose of 15 mg/kg there was a decrease in the frequency of CD 11 expressing TNF-α and it was not significant when compared to the negative control (Figure 4b).
Effect of ARTAVOL® extract on CD335 (Natural Killer Cells) response in P. berghei-Infected mice
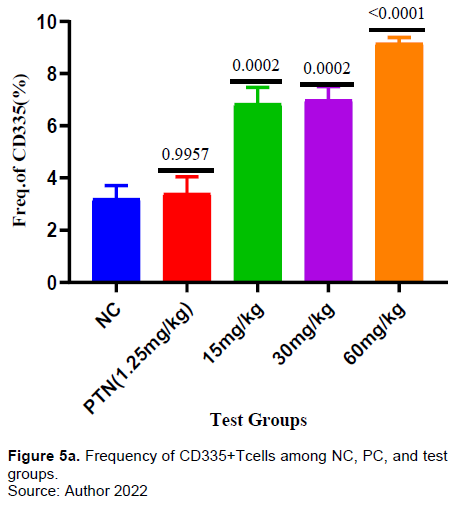
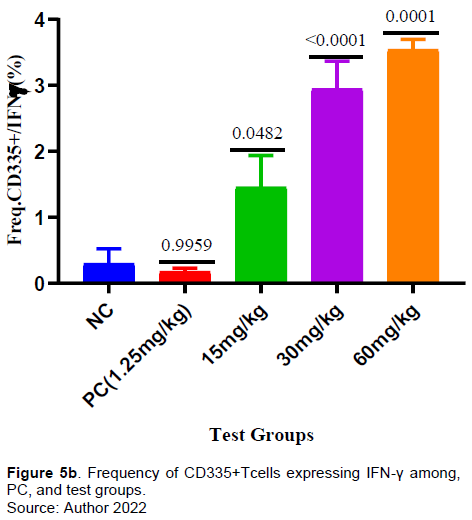
It was observed that a significant increase in the frequency of CD335 in the test groups (15, 30, and 60 mg/kg) when comapred with the negative conrol (Figure 5a). Similarly, there was a significant increase in frequency of CD335 expressing IFN-γ in the all the test groups (15, 30, and 60 mg/kg) when compared with the negative control (Figure 5b).
DISCUSSION
Herbal medicines have been used over the years to treat malaria and have been reported to regulate the immune system in malaria (Afolayan et al., 2020; Kamau, 2022). ARTAVOL® is an herbal product used to prevent malaria in Uganda. Investigations in vivo reported the antimalarial effects of the product (Ogwang et al., 2011b). However, there was no available data on its immunomodulatory activity during P. berghei infection which has been reported in this study.
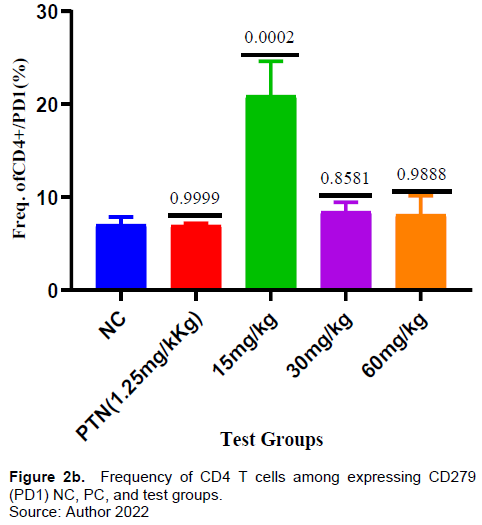
In this study, a dose- dependent was observed from 15 to 60 mg/kg in the frequency of CD4+ T cells (Figure 2a). For instance, at 30 and 60 mg/kg, an insignificant frequency increase was exhibited compared to the negative control which could be from the increase in antagonizing component(s) as the concentration increased. Kurup et al. (2019) reported that drugs may induce T cell exhaustion which can down- regulate T cell function (Kurup et al., 2019). The cell surface expression of the exhaustion marker PD-1(CD279)) was assessed (Figure 2b).
The herbal test doses (30 and 60 mg/kg) did not show significant increase in the frequency of CD4+ T cells expressing PD-1(CD 279). This suggests that the herbal extract in the test doses of 30 and 60 mg/kg did not induce CD4+ T cell proliferation. Konkel et al. (2010) reported that PD-1 provide inhibitory signals that dampen T cell function and restrains their clonal expansion (proliferation) to an immunogenic stimulus (Konkel et al., 2010). There were observable significant activities in the frequency of CD4+ T cells expressing IL-10 (Figure 2c) in all the test doses (15-60 mg/kg) of the extract. A review study by Rajiv Kumar et al., (2019) reported that IL-10 regulates infection mediated inflammation during malaria.
This study observed no significant increase in the frequency of CD8+ T cells (Figure 3a) in all the test doses compared to both positive and negative controls suggesting that the extract did not cause enough proliferation of CD8+T cells. This study’s findings indicated there was a significant increase of CD8+T cells expressing IFN-γ in all extract tests (Figure 3b) compared to the negative control. Similarly, the CD8+ T cells expressing TNF-α increased significantly in the test doses of 30 and 60 mg/kg (Figure 3c). ARTAVOL® contains phytochemicals like coumarins, flavonoids, triterpenes, and sterols among others. Flavonoids and triterpenes have been reported to induce the release of flavonoids, NF-α, and to modify intracellular signaling pathways in immune cells (Afolayan et al., 2020; Brindha, 2016; Mavondo et al., 2019).
In this present study, all the test doses of extract showed a significant increase in the frequency of CD 11(Monocytes) compared to negative control (Figure 4a). Likewise, the extract test doses (30 and 60 mg/kg) induce a significant release of TNF-α by CD 11 compared to the negative control (Figure 4B). Monocytes have been reported to exhibit protection against malaria and reduce parasite burden through cytokine production and phagocytosis (Chua et al., 2013; Dobbs et al., 2020; Ortega-Pajares and Rogerson, 2018).
Furthermore, the extract test doses at 15, 30 and 60 mg/kg showed a significant gradual increase in the frequency of CD 335 (Natural killer cells) compared to the negative control (Figure 5a). Also, in this study there was observable significant increase in production of IFN-γ by CD 335 (Natural killer cells) by all the extract doses compared to the negative control (Figure 5B). Various phytochemicals (e.g., allagic acid, flavonoids, curcumin, and quercetin) have been reported work by either directly affecting the pathogens or indirectly stimulating the innate and adaptive defense mechanisms of the host (Grudzien and Rapak, 2018; Sodagari et al., 2018).
CONCLUSION
The aqueous extract of ARTAVOL® possesses variable immunomodulatory activity and showed ability to increase the frequency of monocytes and natural killer cells. IFN-γ and TNF-α levels showed a significance increase suggesting the extract induced their responses. However, there is need to investigate the effect of the extract antibody mediated immune response.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors would like to thank BEI Resources USA for provision of P. berghei ANKA parasites, PHARMBIOTRAC and Mr. David Nkwangu for Laboratory extraction of ARTAVOL®.
FUNDING
The research was partly funded by African Development Bank (AfDB)–HEST (Ministry of Education and Sports, Uganda) project and Faculty of Medicine, Mbarara University of Science and Technology.
REFERENCES
|
Afolayan FI, Adegbolagun O, Mwikwabe NN, Orwa J, Anumudu CJSA (2020). Cytokine modulation during malaria infections by some medicinal plants. Scientific African 8:e00428. |
|
|
Ajayi CO, Elujoba AA, Okella H, Oloro J, Raymond A, Weisheit A, Tolo CU, Ogwang PE (2020). In vivo antimalarial activities of five Ugandan medicinal plants on Plasmodium berghei in mice. |
|
|
Bedi O, Krishan P (2020). Investigations on acute oral toxicity studies of purpurin by application of OECD guideline 423 in rodents. Naunyn-Schmiedeberg's Archives of Pharmacology 393(4):565-571 |
|
|
Brindha PJIJOGP (2016). Role of phytochemicals as immunomodulatory agents: A review. International Journal of Green Pharmacy 10(1). |
|
|
Chinsembu KCJAT (2015). Plants as antimalarial agents in Sub-Saharan Africa. Acta Tropica 152:32-48. |
|
|
Chua CLL, Brown G , Hamilton JA, Rogerson S, Boeuf PJTIP (2013). Monocytes and macrophages in malaria: protection or pathology? 29(1):26-34.. |
|
|
Dobbs KR, Crabtree JN, Dent AEJIR (2020). Innate immunity to malaria-the role of monocytes. Immunology Review 293(1):8-24.. |
|
|
Ekor MJFIP (2014). The growing use of herbal medicines: issues relating to adverse reactions and challenges in monitoring safety. Frontiers in Neurology 4:177. |
|
|
Greenwood BJMJ (2010). Anti-malarial drugs and the prevention of malaria in the population of malaria endemic areas. Malaria Journal 9(3):1-7. |
|
|
Grudzien M, Rapak AJJOIR (2018). Effect of natural compounds on NK cell activation. Journal of Immunology Research. |
|
|
Kamau LW (2022). Anti-plasmodial and Immunomodulatory Effects of Combined Ajuga remota and Caesalpinia volkensii Leaf Extracts during Malaria Infection in Balb/c Mice. Doctoral dissertation, JKUAT-COHES. |
|
|
Konkel JE, Frommer F, Leech MD, Yagita H, Waisman A, Anderton SMJI (2010). PD?1 signalling in CD4+ T cells restrains their clonal expansion to an immunogenic stimulus, but is not critically required for peptide?induced tolerance. Immunology 130(1):92-102. |
|
|
Kurup SP, Butler NS, Harty JTJNRI (2019). T cell-mediated immunity to malaria. Nature Reviews Immunology 19(7):457-471.. |
|
|
Lorke DJAOT (1983). A new approach to practical acute toxicity testing. Archives of Toxicology 54(4):275-287.. |
|
|
Loue S (2020). Animal Experimentation in Biomedical Science. In Case Studies in Society, Religion, and Bioethics (pp. 209-228). Springer, Cham. |
|
|
Mavondo GA, Mkhwanazi BN, Mzingwane ML, Dangarembizi R, Zambuko B, Moyo O, Musiwaro P, Chikuse FF, Rakabopa C, Mpofu T (2019). Malarial inflammation-driven pathophysiology and its attenuation by triterpene phytotherapeutics. Parasitology and Microbiology Research. IntechOpen. 10.5772/intechopen.88731. |
|
|
Mesa LE, Vasquez D, Lutgen P, Vélez ID, Restrepo AM, Ortiz I, Robledo SMJRDSBDMT (2017). In vitro and in vivo antileishmanial activity of Artemisia annua L. leaf powder and its potential usefulness in the treatment of uncomplicated cutaneous leishmaniasis in humans. Revista da Sociedade Brasileira de Medicina Tropical 50:52-60. |
|
|
Ogwang PE, Ogwal JO, Kasasa S, Ejobi F, Kabasa D, Obua C (2011). Use of Artemisia annua L. infusion for malaria prevention: mode of action and benefits in a Ugandan community. British Journal of Pharmaceutical Research 1(4):124. |
|
|
Ortega-Pajares A, Rogerson SJJFII (2018). The rough guide to monocytes in malaria infection. Frontiers in Immunology. |
|
|
PRISM G (2018). 8.0. 2. |
|
|
Schlagenhauf P, Wilson ME, Petersen E, Mccarthy A, Chen LH (2019). Malaria chemoprophylaxis. Travel medicine. Elsevier. |
|
|
Shellvarajah M, Hatz C, Schlagenhauf PJTM, Disease I (2017). Malaria prevention recommendations for risk groups visiting sub-Saharan Africa: a survey of European expert opinion and international recommendations. Travel Medicine and Infectious Disease 19:49-55. |
|
|
Sodagari HR, Aryan Z, Abdolghaffari AH, Rezaei N, Sahebkar AJJOP (2018). Immunomodulatory and Anti-Inflammatory Phytochemicals for the Treatment of Inflammatory Bowel Disease (IBD)-Turning Strong Rationale into Strong Evidence? 21(4):294-295. |
|
|
Tizifa TA, Kabaghe AN, Mccann RS, Van Den Berg H, Van Vugt M, Phiri KSJCTMR (2018). Prevention efforts for malaria. Current Tropical Medicine Reports 5(1):41-50. |
|
|
Trottier H, Elliott SJJCJOPH (2021). World Health Organization recommends first malaria vaccine. Canadian Journal of Public Health 112(6):967-969. |
|
|
Tugume P, Kakudidi EK, Buyinza M, Namaalwa J, Kamatenesi M, Mucunguzi P, Kalema JJJOE (2016). Ethnobotanical survey of medicinal plant species used by communities around Mabira Central Forest Reserve, Uganda. 12(1):1-28. |
|
|
Willcox M, BODEKER GJB (2004). Traditional herbal medicines for malaria. Bmj 329(7475):1156-1159. |
|
|
World Health Organization (2015). Global technical strategy for malaria 2016-2030, World Health Organization. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


