Review

ABSTRACT
Available therapeutic interventions for managing preterm labour have not been consistently successful due to controversies related to its etiology. Multiple mechanisms, including inflammation play a significant role in the pathogenesis of preterm labour. The connective tissue extracellular matrix of the amniochorion contains collagen fibres that maintain the tensile strength of the amniochorion, resisting mechanical stress and preventing rejection of the fetal allograft. Expression of pro-inflammatory mediators in the amniochorion triggers production of prostaglandins in the uterus and enzymatic degradation of the resilient extracellular matrix of the fetal membranes by matrix metalloproteinases leading to uterine contractions and cervical remodelling resulting in preterm labour. This review appraises the pathophysiological mechanisms of pro-inflammatory mediators in spontaneous preterm labour and their associations with multi-factorial etiological pathways. The physiological pathways and biological mechanisms of uterine activity during pregnancy and parturition are also discussed. Finally, the review provides an overview of the biological basis of common therapeutic agents for treating preterm labour. In this review, keywords related to pathophysiological mechanisms of maternal pro-inflammatory mediators in preterm labour and clinical management were used in the literature search from the PubMed and Google Scholar databases. The snowball sampling methodology was further employed to obtain a comprehensive literature search.
Key words: Preterm labour, pro-inflammatory mediators, pathophysiology.
INTRODUCTION
Preterm labour is defined as the spontaneous onset of uterine contractions resulting in the progressive cervical dilatation or effacement prior to 37 weeks of gestation (Chatterjee et al, 2007; Stabile et al., 2000). Preterm labour may result in preterm birth which is generally classified based on the gestational age at birth as: extremely preterm (before 28 weeks), very preterm (between 28 and 32 weeks) and moderate preterm (after 32 weeks and before 37 weeks) (Blencowe et al., 2013; Soon, 2012). The moderate preterm birth can further be described as late preterm if the onset of it occurs between 34 and 37 weeks of gestation (Blencowe et al., 2013;Soon, 2012). Etiologically, preterm birth can either be spontaneous (natural onset of labour or preterm premature rupture of membranes) or provider-initiated (medically induced labour or caesarean delivery) depending on the clinical presentation (Adu-Bonsaffoh et al., 2019; Souza et al., 2016; Vinturache et al., 2016).
The global incidence of preterm birth is 11% which is equivalent to approximately 15 million babies born preterm each year (Galindo-Sevilla et al., 2019; Perales, 2018). The incidence ranges from 5% among the European countries to nearly 20% in some African countries (Adu-bonsaffoh et al., 2020; Boyle et al., 2017; Perales, 2018). As the principal cause of increased morbidities and mortalities among children less than five years old, prematurity has become a major public health concern (Bayar et al., 2020; Perales, 2018)and compromises the emotional and psychological states of many families with preterm infants born to them. This is attributable to the accelerated economic burdens exerted due to increased cost of health care and severe infant morbidities including learning demands (Adu-Bonsaffoh et al., 2019; Boyle et al., 2017; Purisch and Gyam, 2017). Evidence has shown that infants who are born at extremely low gestational age are at increased risk of severe longstanding health complications (Boyle et al., 2017; Mccormick et al., 2011; Purisch and Gyam, 2017; Rubens et al., 2014). The consequences of preterm birth are common during the neonatal period and may persist into adulthood. Studies have also shown that male infants born preterm have poorer prognosis and are at increased risk of health complications compared to their female counterparts (Boyle et al., 2017; Driscoll et al., 2018; Kent et al., 2011).
The actual aetiology of preterm labour is not known, although multiple mechanisms have been propounded for the pathogenesis of preterm labour. For instance, intrauterine infections, inflammation, mechanical stress or over distension of the uterus, intrauterine growth restriction (IUGR) and uteroplacental hypoxia or haemorrhage are said to trigger the onset of preterm labour (Adu-Bonsaffoh et al., 2020; Boyle et al., 2017; Hamilton et al., 2013; Rubens et al., 2014). Additionally, maternal race (black women, African-American or Afro-Caribbean) or ethnicity, younger maternal age, advanced maternal age (>35 years), cigarette smoking, low maternal weight, obesity, multiple pregnancy, use of assisted reproductive techniques, maternal history of preterm birth and the marginalised may also contribute to the onset of preterm labour (Boyle et al., 2017; Goldenberg et al., 2009). Regardless of the predisposition, maternal pro-inflammatory mediators play a central role in the initiation of preterm labour. In the last decade, the term ’placental inflammation’ has surfaced in clinical practice. Recent evidence indicates that obliteration of the placental structures and impaired function in the maternal-fetal interface leads to obstetric complications (e.g. villitis, chronic chorioamnionitis and chronic deciduitis) and poor pregnancy outcomes such as intrauterine growth restriction (IUGR), preeclampsia, intrauterine fetal demise (IUFD) and preterm labour (Kim et al., 2016; Tanacan et al., 2019; Yang et al., 2019). The pathophysiological mechanisms related to the placental injury and inflammation may be due to infiltration of the placenta by white blood cells (e.g. lymphocytes and macrophages) from the presence of viral, bacterial, parasitic infection, toxic and metabolic, extractable nuclear antigen (ENA), anti-double stranded DNA (anti-dsDNA) positivity and other autoimmune disorders such as systemic lupus erythematosus (SLE). These mechanistic cascades increase the risk of rejection of the fetal allograft and preterm labour (Kim, et al., 2016; Tanacan et al., 2019; Tanacan, 2021).
It is worth emphasizing that accurate prediction of preterm birth has been extremely problematic due to the limited knowledge and multi-factorial aetiologies involved in the pathogenesis of preterm labour (Bas and Tola, 2018). The role of complete blood count (CBC) in predicting the risk of preterm birth has been well established in contemporary times. For instance, CBC inflammatory biomarkers such as white blood cells (WBC), hemoglobin (HGB), platelet distribution width (PDW), neutrophiles, lymphocytes and neutrophil-lymphocyte ratio (NLR) with increased sensitivity and specificity have been found to be positive predictive markers for increased risk of preterm birth compared to term delivery (Bas and Tola, 2018; Ma et al., 2020). To prevent placental inflammation and associated obstetric complications, low-dose low-molecular-weight heparin prophylaxis appears to be effective in preventing metabolic and immunologic disorders leading to placental inflammation (Beksac et al., 2020).
Clinical management of preterm labour is extremely challenging with frequent reports of poor outcomes especially when uterine contractions are established with progressive cervical changes. The unpredictability of the clinical course of preterm labour results from the unresolved pathophysiology of the condition. The major hypothesized pathophysiological mechanisms of preterm labour include inflammation resulting from multiple aetiological pathways. Improved understanding of the mechanistic role of pro-inflammatory mediators in preterm labour is critical in clinical management especially inflammation associated with chorioamnionitis and urinary tract infections (Galindo-Sevilla et al., 2019). The aim of this review was to appraise the pathophysiological mechanism of pro-inflammatory mediators in spontaneous preterm labour and their associations with multi-factorial etiological pathways. The paper also recognises and discusses the pathophysiology of uterine contractions and the phases of uterine smooth muscle activity in the non-pregnant state, during pregnancy and parturition. In this paper, essential physiological pathways and the biological basis of available therapeutic agents (tocolytics) for preterm labour are explored.
MATERIALS AND METHODS
A literature review was done on researches focused on the pathophysiological mechanisms of maternal pro-inflammatory mediators in preterm labour and the management strategies. PubMed and Google scholar were used as medical literature databases for this review. The keywords used for the literature search included: Pathophysiological mechanisms, inflammation, matrix metalloproteinases, spontaneous preterm labour, preterm birth, uterine activity, tocolytics, treatment of preterm labour and placental injury. The snowball search technique was also employed to search for articles on the subject area.
PROBABLE ETIOLOGIES OF PRETERM LABOUR
The role of genetics in the pathogenesis of preterm labour has been duly recognized and well documented. Emerging evidence indicates that preterm birth appears to be transmitted primarily in a matrilineal manner across generations and is greatly influenced by mutual environmental factors. Thus, the risk of a woman having a preterm delivery is heightened if her maternal biological relations had preterm deliveries but appears not to follow paternal lines (Bezold et al., 2013; Wise and Sharma, 2010). In human genomic studies, changes in single nucleotide sequence in genes due to variation in environmental factors were found to be associated with disorders such as polyhydraminos, myotonic dystrophy, cervical incompetence (Adu-bonsaffoh et al., 2020; Behrman and Butler, 2007; Wise and Sharma, 2010)and twin gestation which increase the risk of preterm labour and may be transmitted from one generation to the next (Behrman and Butler, 2007; Ramos et al., 2016). These observed single nucleotide polymorphisms in the gene can respond to inflammatory stimuli such as infection resulting in the production of pro-inflammatory mediators and breakdown of extracellular matrix by matrix metalloproteinases leading to preterm labour (Figure 1) (Ramos et al., 2016). Also, because of the genotypic variations among different individuals, exposure to environmental pollutants put them at increased risk of preterm labour (Table 1) (Adu-bonsaffoh et al., 2020; Behrman and Butler, 2007). Table 1 gives a summary of the probable causes or contributory factors and their biological mechanisms that lead to the onset of preterm labour.labour are explored.
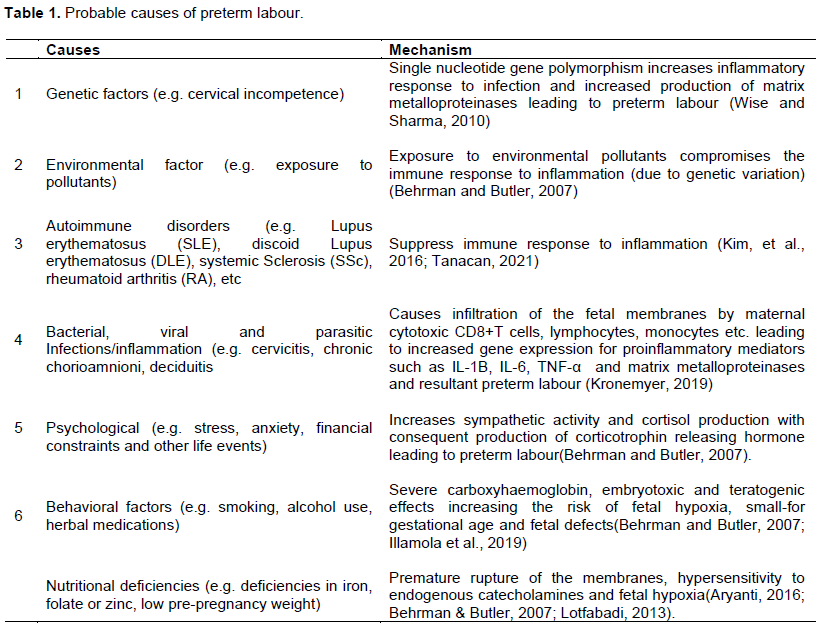
Evidence has shown that maternal psychological stress, anxiety, financial difficulties and other life events increase endogenous catecholamines and cortisol release which stimulate premature production of placental corticotrophin-releasing hormone (CRH) thereby activating the biological cascade leading to the onset of preterm labour (Behrman and Butler, 2007). Psychological stress also increases maternal vulnerability to infection and inflammation due to weakened immune system (Figure 1) (Behrman and Butler, 2007). Behavioural factors such as smoking and alcohol use are associated with increased risk of preterm labour. Smoking and alcohol have teratogenic effects and habitual engagement in these agents during pregnancy puts them at high risk of sudden infant death syndrome (SIDS) due to variability in autonomic nervous system function (Burguet et al., 2004; Cogswell et al., 2003; Fifer et al., 2009). The nicotine and carbon monoxide contained in smoke cause fetal hypoxia due to carboxyhaemoglobin and vasoconstriction (Cogswell et al., 2003). They increase the risk of low birth weight, fetal defects and placental complications and compromise immune response to inflammation leading to preterm premature rupture of membranes and preterm labour (Figure 1) (Behrman and Butler, 2007; Cogswell et al., 2003; Fifer et al., 2009). Consumption of alcohol during pregnancy has also proven to have detrimental effect on fetal growth and development leading to intrauterine growth restriction (IUGR) which poses a potential risk for preterm labour (Table 1 and Figure 1) (Albertsen et al., 2004; Behrman and Butler, 2007).
Furthermore, pervasive indulgence in illegal drugs such as marijuana and cocaine as well as the use of herbal medications in pregnancy have severe embryotoxic and teratogenic effects increasing the risk of small-for gestational age and fetal defect (Illamola et al., 2019; Walker et al., 2000). On the other hand, cocaine causes elevated blood pressure due to vasoconstriction since these psychoactive agents appear to interfere with the reuptake of serotonin and catecholamines leading to compromised immune response to inflammation (Table 1 and Figure 1) (Applová et al., 2018; Behrman and Butler, 2007). Nutritional deficiencies (e.g. deficiencies in iron, folate or zinc, low pre-pregnancy weight) have been implicated in preterm labour. For instance, deficiency in iron causes iron deficiency anaemia which increases the synthesis and hypersensitivity to endogenous catecholamines leading to maternal and fetal hypoxia which, in-turn, stimulates synthesis of fetal CRH and cortisol (Table 1). The increased cortisol production may activate the biological cascade leading to preterm premature rupture of membranes, gestational hypertension, eclampsia IUGR and preterm labour (Allen, 2001; Behrman and Butler, 2007; Zhang et al., 2009). In another pathway, iron deficiency anaemia increases risk of infection and inflammation leading to preterm birth (Allen, 2001; Zhang et al., 2009).
Physiologically, vitamin C stimulates the synthesis of collagen fibres of connective tissue extracellular matrix of the amnionchorion and deficiency in vitamin C depletes collagens leading to preterm premature rupture of membranes and preterm labour (Aryanti, 2016; Behrman and Butler, 2007; Lotfabadi, 2013). Similarly, folate, zinc and pre-pregnancy weight deficiencies increase the risk of restricted fetal growth and birth defects (Figure 1) (Behrman and Butler, 2007; Wang et al., 2015). To buttress the importance of specific nutrients, adequate maternal levels of fat-soluble nutrients provide antioxidative, anti-inflammatory, and immunomodulatory health benefits which are vital in preterm birth prevention. For instance, omega-3 fatty acids facilitate increased production of specialized anti-inflammatory mediators with resultant reduction in preterm birth risk. Combined effects of these nutrients support appropriate placental organogenesis and function (Thoene et al., 2020).
UTERINE SMOOTH MUSCLE ACTIVITY IN THE NON-PREGNANT STATE
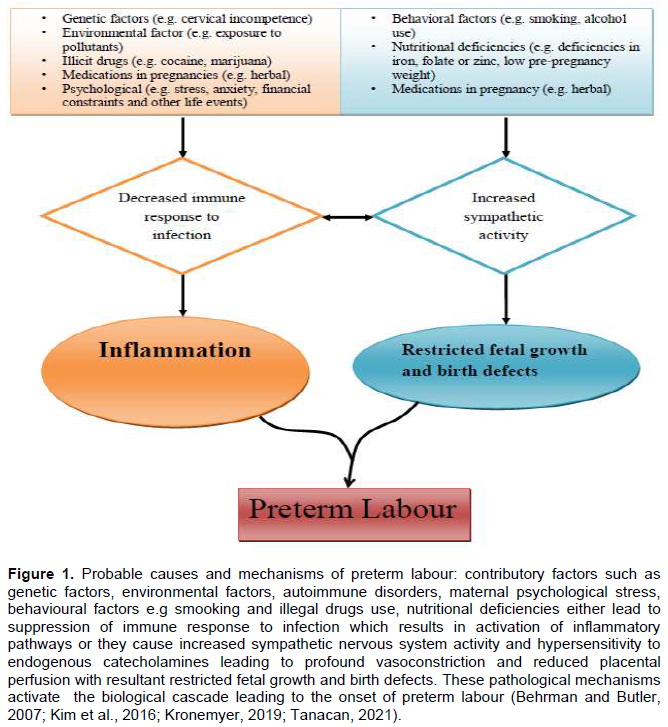
The uterus acts as a temporal receptacle which nurtures and nourishes the implanted blastocyst while thwarting early expulsion of the product of conception throughout pregnancy. It is a contractile smooth muscular organ made up of fibrous connective tissue (Phung et al., 2020)that exhibits spontaneous and rhythmic contraction and relaxation before, during and after pregnancy (Bulletti and De Ziegler, 2005). The non-pregnant uterus is not totally a quiescent organ and exhibits intrinsic contractile properties. However, contractions in the non-pregnant uterus vary physiologically from the uterine contractile activities during pregnancy (Sanborn, 2001). Accordingly, both non-invasive intrauterine pressure (IUP) and invasive ultrasound (US) scan recordings have demonstrated that the non-pregnant uterus undergoes patterns of phasic wave-like contractions throughout the menstrual cycle (Bulletti and De Ziegler, 2005; Fanchin and Ayoubi, 2009; Sammali et al., 2018)which are governed by ovarian steroid hormones (that is, oestrogen and progesterone) (Bulletti and De Ziegler, 2005). Within the subendometrial layer of the myometrium, cyclic patterns of oestrogen and progesterone receptors are expressed which modulate the activity of the non-pregnant uterus(Bulletti and De Ziegler, 2005; Gestel et al., 2003). These wave-like contractile patterns of the non-pregnant human uterus vary depending on the phase of the menstrual cycle (Figure 2) (Fanchin and Ayoubi, 2009; Gestel et al., 2003). The rhythmic uterine contractions in the non-pregnant uterus favours sperm transport and oocyte migration in the fallopian tubes, fertilization and embryonic transport from the tubes into the uterine cavity to facilitate embryonic nidation and implantation to occur (Bulletti et al., 2000; Fanchin and Ayoubi, 2009; Gestel et al., 2003; Sammali et al., 2018). Disruptions in this physiological uterine smooth muscle property are associated with disorders such as dysmenorrhoea, spontaneous and recurrent abortions, endometriosis, implantation failures and infertility (Fanchin and Ayoubi, 2009; Gestel et al., 2003).
During menstruation or early follicular phase, the uterus exhibits primarily antegrade (from fundus to cervix) labour-like and expulsive contractions (Figure 2) involving all layers of the myometrium to evacuate the content of the uterus (menses). Uterine contractions during menstruation are often felt and can be associated with blunt pain (dysmenorrhoea) requiring medications if contractions are vigorous (Gestel et al., 2003; Ziegler et al., 2001). This period of luteofollicular changeover is largely under the influence of progesterone depletion due to spontaneous degeneration of the corpus luteum and increased gene expression for prostaglandins by uterine tissues (Bulletti et al., 2000; Ziegler et al., 2001). In both mid and late follicular phase of the menstrual cycle, only the subendometrial layer exhibits a progressive increased in wave-like uterine contraction patterns which are retrograde (from cervix to fundus) (Figure 2). This retrograde uterine contractions, often not perceived by the woman, coupled with proliferation of endometrial glands which aid the transport of sperm towards the distal end of the fallopian tubes where fertilization takes place and normally terminates at the pre-ovulatory period (Fanchin and Ayoubi, 2009; Gestel et al., 2003; Rabotti et al., 2015; Sammali et al., 2018; Ziegler et al., 2001). These wave-like contractions and glandular proliferation during the follicular phase are controlled by increased estrogen (E2) levels (Figure 2) (Fanchin and Ayoubi, 2009; Gestel et al., 2003; Rabotti et al., 2015; Sammali et al., 2018)indicative of oestrogen predominance in the proliferative phase of the uterine cycle.
The post-ovulation period is characterized by progressive uterine quiescence facilitated by significant rise in progesterone levels following the successful development to differentiate into the fetal membranes comprising rich in connective tissues in both the amnion and the chorion (Vadillo-ortega and Estrada-gutie, 2005). The connective tissue extracellular matrix of the amniochorion contains collagen fibres that maintain the tensile strength of the amniochorion; resisting mechanical stress and prevents rejection of the fetal allograft. Specific collagen fibres of the fibrous connective tissue within the amniotic compact layer provide the tensile strength while collagen fibres of both reticular and spongy layers of the chorion provide mechanical support (Vadillo-ortega and Estrada-gutie, 2005). The major types of collagen fibres include: Type I, II, III, IV, V and VI collagens which are embedded in the fibrous tissue (Vadillo-ortega and Estrada-gutie, 2005).
Typically, the inner layer of the placenta is made up of the amniontic membrane which is composed of amnion and chorion. The amnion has five separate layers; the epithelium, basement membrane, compact layer, fibroblast layer and spongy layer (Koob et al., 2014; Malhotra and Jain, 2014). The epithelium which consists of a single layer of epithelial cells is proximal to the developing. The basement membrane of the amnion is a thin layer composed of collagens III and IV and noncollagenous glycoproteins laminin, nidogen, and fibronectin. The compact layer is dense nearly without cells and forms the main fibrous structure of the amnion. The fibroblast layer of the amnion is the thickest and consists of fibroblasts embedded in a loose collagen network with abundance of noncollagenous glycoproteins. The outermost spongy layer forms the interface between the amnion and chorion and composed of a nonfibrillar meshwork of collagen III and an abundant content of proteoglycans and glycoproteins (Koob et al., 2014; Malhotra and Jain, 2014). The chorion is made up of a reticular layer, basement membrane and trophoblast layer which is adhered to the maternal decidua. The reticular layer contacts the spongy layer of the amnion and forms a majority of chorion’s thickness. The reticular network is composed of collagens I, III, IV, V, and VI. The basement membrane anchors the trophoblasts to the reticular layer with collagen IV, fibronectin, and laminin.
The trophoblast layer is the deepest layer which attaches to the decidua (Koob et al., 2014). Type I and III collagen fibres are known to provide tissue support while type II, IV, V and VI provide scaffoldings in maintaining tensile strength(Vadillo-ortega and Estrada-gutie, 2005). Distortion in the nomenclature of these collagen fibres diminishes the tensile strength of the amniochorionic extracellular matrix which increases myometrial activity and cervical remodelling leading to the onset of preterm labour (Vadillo-ortega and Estrada-gutie, 2005).
PHYSIOLOGICAL UTERINE ACTIVITY DURING PREGNANCY AND PARTURITION
During pregnancy and labour, the myometrium undergoes distinct molecular changes from noncontractile phynotype (quiescence) to contractile phenotype depending on the gestational age (Phung et al., 2020; Vannuccini et al., 2016; You et al., 2011). This process of uterine activity during pregnancy and parturition can be divided into at least four separate phases (Challis et al., 2002; Felice et al., 2010; Schwarz and Page, 2003). The four recognized phases of uterine activity include quiescence, activation, stimulation and involution (Figure 2).
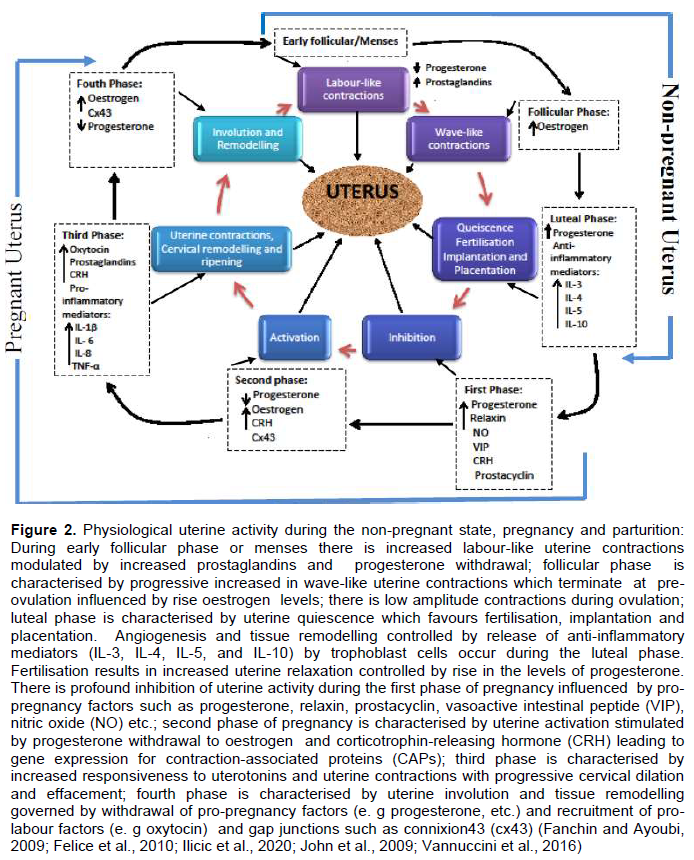
In the first phase (uterine quiescence) which occurs in pregnancy, there is increased inhibition of uterine activity by either separate or combined autocrine-paracrine signalling transduction stimulated by pro-pregnancy factors such as progesterone, prostacyclin (PGI2), relaxin, parathyroid hormone-related peptide (PTHrP), calcitonin gene-related peptide, adrenomedullin, vasoactive intestinal peptide (VIP), nitric oxide, and CRH, which maintains the uterus in a relatively quiescent state (Challis et al., 2002; Ilicic et al., 2020; Schwarz and Page, 2003). This allows angiogenesis and tissue remodelling especially around the placentation site to improve adequate placental circulation, fetal nutrition and intrauterine growth (Figure 2) (Shynlova et al., 2013). Alteration in the production of these agents during late gestation may lead to the onset of preterm or term labour, whereas administration of these compounds or their analogues may help maintain uterine quiescence. Uterine smooth muscle relaxants act by signalling through binding and activation of G-protein stimulatory (Gαs) subunit of the G-protein coupled receptor (GPCR) located on the surface of the myocytes. Activated GTP bound Gαs activates adenylate cyclase or guanylate cyclase causing increased intracellular concentrations of cyclic adenosine monophosphate (cAMP) or cyclic guanosine monophosphate (cGMP). These nucleotides interfere with intracellular calcium release and reduce the activity of myosin light chain kinase (MLCK) which are required for shortening of the myofilaments and smooth muscle contractions (Figure 4) (Challis et al., 2002; Ilicic et al., 2020; Schwarz and Page, 2003).
As pregnancy approaches term, the myometrium transitions from its quiescence state to activation. This second phase of uterine activity is characterised by progressive progesterone diminution and transtition to oestrogen and CRH dominance associated with increased mechanical stretch or uterotrophic sensitisation (Figure 2) (Ilicic et al., 2020; Schwarz and Page, 2003; Vannuccini et al., 2016). This leads to increased expression of contraction-associated proteins (CAPs) including connexion 43 (Cx43, a key component of gap junctions), agonist receptors (prostaglandins and oxytocin) as well as increased influx of calcium ions into the uterine myocytes (Challis et al., 2002; Ilicic et al., 2020; Schwarz and Page, 2003; Vannuccini et al., 2016). During the second phase of uterine activity, upregulation of GJA1/Cx43 gene mediate hypersensitivity of receptors to uterotonic agonists such as PGs and oxytocin which generate high intensity and coordinated phasic uterine contractions(Ilicic et al., 2020; Nadeem et al., 2016). Connexion 43 is a gap junction channel formed from hexamers of connexion proteins that plays a critical role in cell to cell coupling and generates synchronous myometrial contractions (Dunk et al., 2012; Nadeem et al., 2016). It allows direct exchange of macro molecules, ions, and second messengers such as cAMP, cGMP, inositol phosphate and Ca2+ between cells which enable channels control and coordinate cellular activity (Figure 4) (Dunk et al., 2012).
Stimulation represents the third phase of uterine activity during parturition and is typified by increased uterine smooth muscle tone stimulated by uterotonins such as PGs, oxytocin, and CRH. This phase is characterised by increased synthesis of pro-inflammatory mediators (that is, IL-1β, IL-8, TNF-α etc.), prostaglandins and white cells (e.g. monocytes and neutrophils) infiltration of the uterus, fetal membranes and cervix; activating a biochemical process that triggers inflammatory reactions within the uterus. Production of pro-inflammatory mediators causes release of matrix metalloproteinases which breakdown the extracellular matrix and increased gene expression for prostaglandins in uterine tissues (Figure 2). This leads to uterine contractions, cervical remodelling and ripening (Gibb et al., 2006; Terzidou, 2007; Vannuccini et al., 2016). Increased expression of prostaglandins receptors and gap junctions also occurs in the third phase of parturition. Prostaglandins stimulate functional fundal contractions, lower segmental contraction of the uterus and facilitate cervical ripening while gap junctions permit cell to cell communication by allowing intercellular exchange of macro molecules such as calcium, cAMPs and cGMPs (Figure 4)(Gibb et al., 2006; Terzidou, 2007).
The last phase of uterine activity is characterised by massive uterine involution to resemble the pregnancy stage and tissue remodelling after delivery of the and placenta, and has been attributed primarily to the release of neuroendocrine oxytocin (Challis et al., 2002; Gibb et al., 2006; Ilicic et al., 2020; Schwarz and Page, 2003; Vannuccini et al., 2016). During this phase, there is a rapid withdrawal of pro-pregnancy factors (e.g. progesterone) that maintains uterine quiescence and increased recruitment of pro-labour factors (e.g. gap junctions, calcium-sensitive potassium channels, and connexions) that stimulate the biochemical changes similar to those occurring in labour (Figure 2) (Terzidou, 2007).
PRO-INFLAMMATORY MEDIATORS AND SPONTANEOUS PRETERM LABOUR
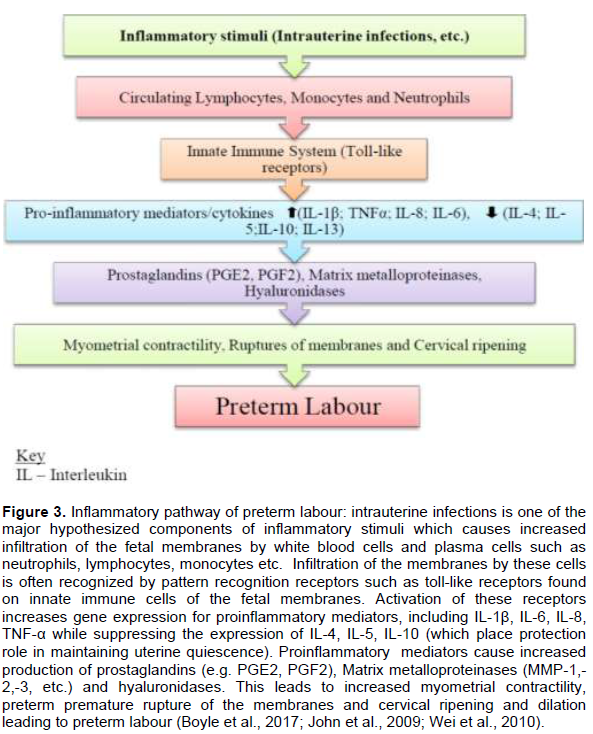
Inflammation plays a crucial role in the process of parturition (Boyle et al., 2017; John et al., 2009). In preterm labour, pathological mechanisms involving the cervix, fetal membranes, placenta, and myometrium activate one or more of the components of the parturition pathways prematurely (John et al., 2009). During preterm labour, uterine activity is changed from a state of quiescence to a pro-inflammatory milieu in a three-step process characterized by uterine contractility, cervical remodelling and membrane activation/rupture which is modulated by pro-inflammatory mediators/cytokines (Boyle et al., 2017; John et al., 2009; Wei et al., 2010). Intrauterine infection is one of the most frequent conditions leading to preterm labour and the pathogenic mechanisms are related to activation of receptors (toll-like receptors) on the innate immune system (John et al., 2009). The significant components of the innate immune system are pattern recognition receptors. Toll-like receptors (TLRs) are a class of pattern recognition receptors that detect pathogen-associated molecular patterns (PAMPS) derived from microorganisms and damage-associated molecular patterns (DAMPS) released from immune cells, stressed and dying cells. These stimulate intracellular signalling cascades leading to the expression of pro-inflammatory mediators/ cytokines by innate immune cells resulting in normal or preterm labour (Figure 3) (Green and Arck, 2020; Silva et al., 2010).
Pro-inflammatory mediators are structurally classified into 4 groups: The 4α helix family members - interleukin 2 (IL-2), interferon gamma (IFN-γ) and IL-10; IL-1 family; IL-17 family; and chemokines. Functionally, these pro-inflammatory mediators/cytokines are grouped into T helper type 1 (Th1) cell reactions (cell-mediated immunity) and T helper type 2 (Th2) reactions (humoral immunity) (John et al., 2009). Inflammatory cytokines are produced by T Lymphocytes (CD4+) which modulate immune response to inflammatory stimuli. Primarily, Th1 cells produce IL-1, IL-2, IL-6, IL-12, IL-15, IL-18, IFN-γ, and tumor necrosis factor alpha (TNF-α) and Th2-cells produce IL-4, IL-5, IL-10, IL-13, and granulocyte macrophage colony stimulating factor (GM-CSF) (John et al., 2009; Noakes and Michaelis, 2013).
The presence of intrauterine infection causes infiltration of circulating monocytes and neutrophils into the myometrium and cervix leading to significant gene expression for interleukins (IL-1β, IL-6, IL-8) and tumour necrosis factor alpha (TNF-α) (Boyle et al., 2017; John et al., 2009). Myometrial contractions are stimulated by the increase of IL-1 and TNF-α which promote influx of calcium into myometrial smooth muscle cells (Boyle et al., 2017; Hutchinson et al., 2014). Pro-inflammatory mediators also cause increased gene expression for prostaglandin synthase coupled with cyclooxygenase-2 gene expression leading to up-regulation of prostaglandins and collagenases in uterine tissues and cervix (Silva et al., 2010). Prostaglandins, PGF2 and PGE2, are also involved in the stimulation of myometrial contractions and cervical ripening (Bakker et al., 2017; Olson and Ammann, 2015; Ousey and Fowden, 2012). Cervical shortening and softening is due to the progressive increased in the amount of hyaluronidase and other enzymes that break down the glycosaminoglycans (GAGS) while collagenases break down the cervical collagen. This leads to increased water absorption by the cervix. IL-4, IL-5, IL-10 and IL-13 produced by Th2-cells play a rather potential protective role in the feto-maternal relationship and prevent preterm labour (Figure 3) (John et al., 2009).
In early stage of inflammation, activation and maturation of neutrophils and macrophages occur by stimulatory effect of pro-inflammatory cytokine such as IL-6 in addition to increased differentiation of natural killer cells resulting in invasion of the cervical and endometrial tissue with subsequent stimulatory effect on the uterine contractility (Osman et al., 2003). Similarly, IL-6 causes increased oxytocin receptors expression on myometrial cells with further enhancement of their responsiveness to oxytocin. In another pathway, IL-6 causes increased production of prostaglandins from activation of the hypothalamic-pituitary-adrenal axis and this result in initiation of uterine contraction and progressive cervical dilatation (Farina and Winkelman, 2005).
Pro-inflammatory mediators also stimulate matrix metalloproteinases (MMPs) such as collagenase, gelatinase, stromelysin, hyaluronidases and prostaglandins expression which promote extracellular matrix degradation, uterine contractions and cervical ripening leading to preterm labour (Figure 3) (John et al., 2009; Kota et al., 2013; Larsen and Hwang, 2011; Vadillo-ortega and Estrada-gutie, 2005). Major proteinases associated with this mechanism are collagenases: 1, 2, 3 and 4, (also known as MMP-1, MMP-8, MMP-13 and MMP-18, respectively); gelatinases A and B (MMP-2 and MMP-9, respectively); and stromelysin-1 (MMP-3) and stromelysin-2 (MMP-10) are normally expressed during labour (Visse and Nagase, 2003; Vadillo-ortega and Estrada-gutie, 2005).
PHYSIOLOGICAL PATHWAYS AND BIOLOGICAL MECHANISMS OF UTERINE CONTRACTION
Series of physiological events occur before, during and after pregnancy which modulate myometrial contractility throughout the menstrual cycle in the non-pregnant uterus, maintenance of pregnancy, promotion of childbirth and enhance involution (Aguilar and Mitchell, 2010; Kota et al., 2013; Sanborn et al., 2005; Sanborn, 2001). These events may include both maternal and fetal characteristics which generate signalling molecules necessary for the stimulation of myometrial contractions during labour (Aguilar and Mitchell, 2010; Kota et al., 2013). Knowledge about the biological mechanisms and pathways that control myometrial contraction and relaxation and how these pathways can be regulated is paramount for clinical practice. Conventional trials in animal models showed that parturition is determined by activation of the fetal hypothalamic-pituitary adrenal (HPA) axis with increased fetal cortisol secretion. Following mechanical stress, activation of HPA pathway leads to reduction in maternal progesterone levels and increased levels of oestradiol. This endocrine imbalance promotes increased intrauterine production of prostaglandins, cervical softening and the onset of myometrial contractions (Bernal, 2003; Phillips et al., 2014).
The contractile state of the myometrium is determined by the interaction between the two major muscle proteins, actin and myosin. This actin-myosin interaction is influenced by myometrial signalling pathways which are broadly categorised into signalling cascades regulating intracellular calcium (Ca2+) concentration and those controlling the contractile apparatus itself (Sanborn et al., 2005; Sanborn, 2001). Abundant in the plasma membrane of the uterine myocytes are L-type calcium ion (Ca2+) channels which are ubiquitous, large conductance, voltage-gated calcium channels (VGCC) (Aguilar and Mitchell, 2010; Pierce, 2010). Binding of an agonist (e.g. oxytocin) to specific receptor causes depolarisation of the myocyte’s membrane potential and opening of the L-type calcium channels leading to rapid influx of extracellular calcium ions and dramatic rise in intracellular calcium ion concentration(Aguilar and Mitchell, 2010; Kota et al., 2013). The plasma membrane of the myocyte also contain other types of calcium channels (that is, T-type calcium channels) which exhibit faster kinetics and greater conductance than the L-type (Figure 4) (Aguilar and Mitchell, 2010).
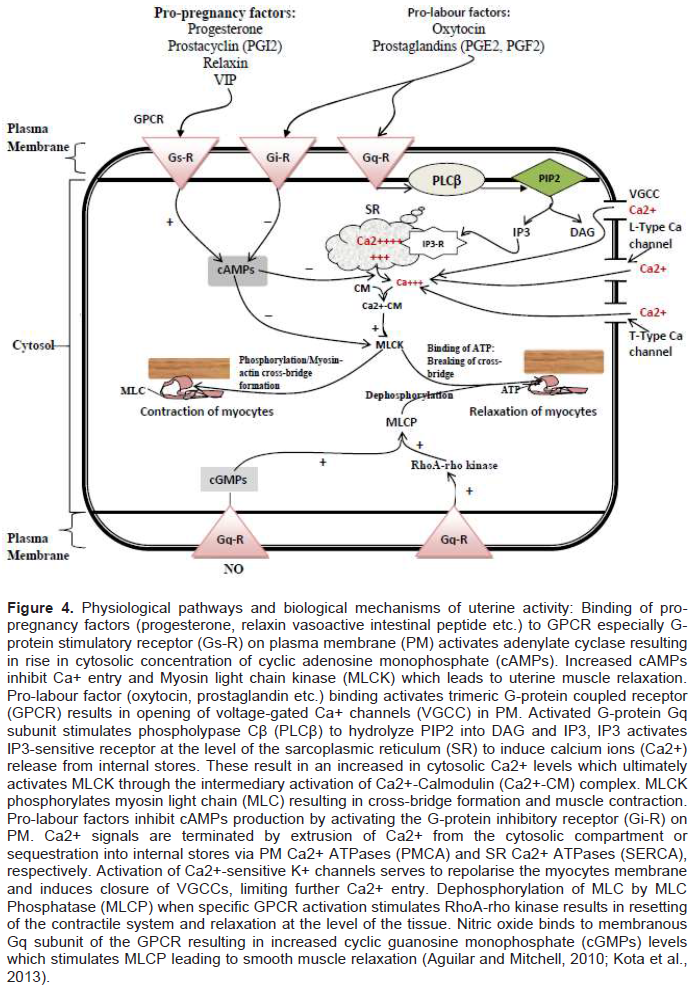
Within the myometrium, agonist interaction with GPCR on the plasma membrane (PM) of myocytes leads to activation of the Gαq subunit of the trimeric G-protein. Activated Gαq subunit also binds and activates membrane-bound phospholypase Cβ which hydrolyzes phosphatidylinositol bisphosphate (PIP2) into inositol-triphosphate (IP3) and diacylglycerol (DAG) (Aguilar and Mitchell, 2010; Kota et al., 2013; Sanborn et al., 2005). IP3 interact with IP3-sensitive receptors (IP3-Rs) on sarcoplasmic reticulum (SR) which causes release of calcium from its storage sites in the SR into the cytosol. Increased cytosolic calcium concentration also stimulates the rynodine receptors to cause Ca2+-induced Ca2+ release (CICR) (Aguilar and Mitchell, 2010; Kota et al., 2013; Sanborn, 2001). Another mechanism, called store-operated Ca2+ entry (SOCE), also regulates Ca2+ flux. When the intracellular Ca2+ stores in the SR are exhausted, SOCE stimulates the PM to permit influx of extracellular Ca2+ into the cytosol (Aguilar and Mitchell, 2010). Increased concentration of Ca2+ in the cytosol leads to binding of calcium to calcium-sensitive protein, calmodulin. The calcium-calmodulin complex activates the enzyme Myosin Light Chain Kinase (MLCK) which in turn causes increased phosphorylation of Myosin Light Chain (MLC) leading to actin-myosin cross-bridge formation and activation of the contractile machinery (Figure 4) (Aguilar and Mitchell, 2010; Kota et al., 2013; Sanborn, 2001).
Relaxation of uterine smooth muscles occurs with removal of cytosolic Ca2+ through closure of PM L-type Ca2+ channels, efflux of Ca2+ into the extracellular compartment through Ca2+-ATPase pumps on plasma membrane and into intracellular Ca2+ stores in SR via SR/ER Ca2+-ATPase (SERCA) pumps (Aguilar and Mitchell, 2010; Kota et al., 2013; Sanborn et al., 2005; Sanborn, 2001). Also, Myosin Light Chain Phosphatase (MLCP) causes dephosphorylation of the myosin light chain which is regulated by signalling pathway through the small G-protein rhoA–rho-associated kinase and protein kinase C (PKC) pathways. Dephosphorylation of the myosin light chain inhibits actin-myosin cross-bridge formation leading to smooth muscle relaxation (Figure 4) (Aguilar and Mitchell, 2010).
Conversely, in postterm pregnancy although the actual aetiology is not yet known, genetics and maternal and fetal factors are implicated in its pathogenesis (Galal et al., 2012; Kota et al., 2013; Mandruzzato et al., 2010). Unlike preterm labour, fetal hypothalamic pituitary adrenal (HPA) insufficiency and a disorder in placental sulphatase activity (an X-linked recessive gene disorder) result in reduced production of estriol (E3) which plays a fundamental role in the pathogenesis of postterm pregnancy (Galal et al., 2012; Mandruzzato et al., 2010). Subsequently, placental CRH production declines with diminution in the positive feedback mechanism on the production of fetal adrenal dehydroepiandrosterone (DHEA). Decreased production of fetal estriol and cortisol ensue which interferes with the biological pathways for spontaneous onset of labour (Galal et al., 2012).
BIOLOGICAL BASIS OF THERAPEUTIC AGENTS FOR PRETERM LABOUR
In recent times, the prediction, prevention and interventions to reduce the incidence of preterm births has remained a major focus for contemporary research in obstetrics (Dickinson et al., 2014; Driul et al., 2014; Newnham, 2017). Several interventions and strategies for the prevention and clinical management of preterm labour have been suggested. These interventions include: Strategies to reduce spontaneous early births, maternal progesterone supplementation, cervical cerclage, minimising maternal exposure to substance abuse and cigarette smoking during pregnancy, cautious patronage of fertility treatments, novel strategies for preventing intrauterine infections, improvement in maternal nutrition and lifestyle modification to reduce maternal stress, adherence to routine antenatal and preterm birth prevention clinics and diagnostic modalities (Dickinson et al., 2014; Mol et al., 2016; Newnham, 2017; Whitaker et al., 2019).
Generally, the use of tocolytics has become the fundamental pharmacological therapy in the clinical management of preterm labour (Haram et al., 2014; Whitaker et al., 2019). The biological basis for the use of tocolytics is to delay delivery for at least 48 hours to allow for corticosteroids and magnesium sulphate administration to enhance fetal lung maturation and prevention cerebral palsy respectively and transfer to a tertiary healthcare facility with neonatal intensive care facilities (Driul et al., 2014; Haram et al., 2014; Whitaker et al., 2019).
Proinflammatory cytokines such as IL-1β, IL-6, and IL-8 have been strongly associated with cervical dilation in preterm labour and medications that decrease the concentration of these mediators constitutes potential preventive interventions (Farina and Winkelman, 2005). The common therapeutic agents which are currently used to abort uterine contractions or maintain uterine quiescence include beta adrenergic receptor agonists (ritodrine, isoxsuprine, salbutamol, terbutaline), prostaglandin synthase inbibitors [including NSAIDS (e.g Indomethacin) and COX-2 inhibitors (e.g Celecoxib)], oxytocin receptor antagonists (Atosiban), calcium channel blockers (Nifedipine) and Magnesium sulphate (Vogel et al., 2014; Whitaker et al., 2019). Tocolytics are usually administered as a monotherapy although a combination of more than one agent maybe required in some cases. However, a recent systematic review by Vogel et al. (2014) did not support the use of a polytherapy of tocolytics over monotherapy and this is partly due to lack of trials of combination regimens of commonly used tocolytics (Vogel et al., 2014).
Beta agonists (betamimetics) bind to beta 2 adrenergic receptors on the surface membranous myometrial smooth muscle cells which activate adenyl cyclase leading to increased intracellular levels of cAMPs and activation of protein kinase A. Increased cAMPs levels decrease intracellular calcium, which specifically suppresses spontaneous and oxytocin-induced uterine contractions. Three different subtypes of beta adrenergic receptors are located in the uterus and beta 2 adrenergic receptors constitutes approximately 80% (Arrowsmith et al., 2010). However, the use agonists of beta 2 adrenergic receptors is associated significant maternal side effects such as palpitations, chest pain, breastlessness and pulmonary oedema in severe cases which limits their clinical use for extended duration (Arrowsmith et al., 2010).
Atosiban is a nona-peptide, desamino-oxytocin analogue and a competitive vasopressin/oxytocin receptor antagonist (Kashanian et al., 2005). Atosiban blocks oxytocin receptors and prevents oxytocin-induced hydrolysis of phosphatidylinositol 4,5-bisphosphate (PIP2) to IP3 and calcium efflux into the cytoplasm (Haas et al., 2014). Thus, it inhibits signalling transduction on the uterine smooth muscle leading to uterine relaxation (Driul et al., 2014; Sanu and Lamont, 2010). Atosiban has minimal side effects with higher safety profile as compared to the other tocolytics(Kosinski et al., 2019).
Selective COX-2 inhibitors such as celecoxib and non-specific COX inhibitors such as indomethacin which are prostaglandin synthase-2 inhibitors inhibit the synthesis of prostaglandins (e.g. PGF2α and PGE2) but do not affect the production of pro-inflammatory mediators (e.g. IL-1β, IL-6, and TNFα) because the action of pro-inflammatory mediators is upstream of COX-2 expression (Loudon et al., 2003; Sakai et al., 2001). Prostaglandins induce uterine contraction via facilitation of myometrial gap junction formation and increasing the concentration of intracellular calcium concentration. Therefore, prostaglandin synthase inhibitors are considered effective tocolytics. However, their usage is limited because they cause premature constriction of the ductus arteriosus, inhibit platelets aggregation and impair renal function and decreased urine production resulting in oligohydramnios. Therefore, the duration of indomethacin use should be restricted and limited to gestational age of < 32 weeks (Hubinont and Debieve, 2011).
Magnesium sulphate is used broadly as a tocolytic and proven to be more effective in fetal neuroprotection and preventing cerebral palsy (Neil and Volpe, 2018). Magnesium sulphate competitively blocks intracellular calcium influx and activation of myosin light chain kinase leading to decreased myometrial contractility. It also inhibits acetylcholine release by competing with calcium at the motor end plate of the neuromuscular junction. Perhaps, its usage is not primarily intended to delay delivery but to prevent cerebral palsy via neuroprotection (Kashanian et al., 2011; Neil and Volpe, 2018). The common side effects include flushing, respiratory depression and cardiac arrest (Haas et al., 2014). Based on our review, all the tocolytic agents in recent use appear to act in areas that interfere with either calcium entry or prostaglandin production which are downstream of pro-inflammatory mediators’ production.
Relatedly, exogenous progesterone supplementation (e.g. 17-α hydroxyprogesterone caproate) in pregnancy is recommended for the prevention of spontaneous preterm delivery especially in women with significant risk factors. Although the use of progesterones is not effective in the treatment of preterm labour, significant successes have been detected in the maintenance of pregnancy and preventing recurrent preterm labour (Ibrahim et al., 2010; Norwitz and Caughey, 2011; Stelzl et al., 2019). Progesterone is a hormone that enhances uterine growth and prevents uterine contractions during pregnancy (Ha et al., 2007; Kota et al., 2013)and has significant physiological effect of maintaining uterine quiescence mediated. In one study, prophylactic treatment with 17-hydroxyprogesterone caproate resulted in approximately 34% reduction in recurrent preterm birth (Caritis et al., 2014; Meis et al., 2003). However, recent evidence on effectiveness of 17-hydroxyprogesterone preterm labour remains conflicting. More recently,
Blackwell et al determined no significant reduction in recurrent preterm birth and no association with increased intrauterine fetal death or early neonatal death with prophylactic use of 17-hydroxyprogesterone in women with a history of preterm birth (Blackwell et al., 2020).
Similarly, the use of nitric oxide (NO) donors such as Nitroglycerine has also been recognized for the treatment and prevention of preterm labour and birth although their clinical usefulness remains conflicting. Due to their potency in mediating uterine relaxation and cervical ripening, NO donors have shown promising results in clinical practice (Duckitt et al., 2014). NO donors are associated with improved placental perfusion by reducing systemic resistance on uteroplacental vessels with significant protective role on the endometruim (Schleussner et al., 2014). Currently, there is insufficient evidence to support the routine use of nitric oxide donors in treating threatened preterm labour (Duckitt et al., 2014).
Further research is recommended concerning the effectiveness of the available interventions for prevention and treatment of preterm labour. In this regard, pathophysiological mechanisms underlying proinflammatory pathway remain central to the clinical management of preterm labour due to the multifactorial etiopathogenesis.
CONCLUSION
Pro-inflammatory mediators play a central role in the pathophysiology of preterm labour and may constitute the main targets for therapeutic interventions in the prevention and clinical management of preterm labour. Imbalances in the pro- and anti-inflammatory pathways may be responsible for preterm labour and postterm pregnancies. Increased expression of pro-inflammatory mediators such as IL-1β, IL-6, IL-8 and TNF-α are associated with increased risk of preterm birth. Although the aetiology of preterm labour remains elusive, alteration in multiple maternal and fetal physiological mechanisms are implicated including genetic predisposition, psychological and environmental influences.
Available medical therapies for preterm labour have not been consistently effective in aborting uterine contractions during preterm labour due to its ill-defined pathophysiology. However, therapeutic interventions primarily targeting the pro-inflammatory pathway seem promising as inflammation plays a central role in the pathogenesis. In-depth understanding of the mechanistic role of pro-inflammatory mediators remains vital to the development of more efficacious therapies and interventions for prevention and treatment of preterm labour. Further research is recommended to provide adequate understanding of the pathophysiological mechanisms of maternal pro-inflammatory mediators in preterm labour.
CONFLICT OF INTERESTS
The authors of this article declare that, no conflict of interest exists.
REFERENCES
|
Adu-Bonsaffoh K, Gyamfi-Bannerman C, Oppong SA, Seffah JD. (2019). Determinants and outcomes of preterm births at a tertiary hospital in Ghana. Placenta 79(1):62-67. |
|
|
Adu-bonsaffoh K, Oppong SA, Dassah ET, Seffah JD (2020). Challenges in preterm birth research: Ghanaian perspective. Placenta 98:24-28. |
|
|
Aguilar HN, Mitchell BF (2010). Physiological pathways and molecular mechanisms regulating uterine contractility 16(6):725-744. |
|
|
Albertsen K, Andersen AMN, Olsen J, Grønbæk M (2004). Alcohol Consumption during Pregnancy and the Risk of Preterm Delivery. American Journal of Epidemiology 159(2):155-161. |
|
|
Allen LH (2001). Biological Mechanisms That Might Underlie Iron ' s Effects on Fetal Growth. American Society for Nutritional Sciences, 131(2):581-589. |
|
|
Applová L, Remiao F, Varner KJ, Pourová J, Costa VM, Št M, Vopršalová M (2018). Comprehensive review of cardiovascular toxicity remysl Mlad enka. Wiley, August 2017, pp. 1332-1403. |
|
|
Arrowsmith S, Kendrick A, Wray S (2010). Drugs acting on the pregnant uterus. Obstetrics, Gynaecology and Reproductive Medicine 20(8):241-247. |
|
|
Aryanti C (2016). Is vitamin C able to prevent premature rupture of membranes? International Journal of Reproduction, Contraception, Obstetrics and Gynecology 5(1):13-16. |
|
|
Bakker R, Pierce S, Myers D (2017). The role of prostaglandins E1 and E2, dinoprostone, and misoprostol in cervical ripening and the induction of labor: a mechanistic approach. Archives of gynecology and obstetrics 296(2):167-179. |
|
|
Bas FY, Tola EN (2018). The role of complete blood inflammation markers in the prediction of spontaneous abortion. Pakistan Journal of Medical Sciences 34(6):1381-1385. |
|
|
Bayar E, Bennett PR, Chan D, Sykes L, MacIntyre DA (2020). The pregnancy microbiome and preterm birth. Seminars in Immunopathology 42(4):487-499. |
|
|
Behrman RE, Butler AS (2007). Preterm birth: causes, consequences, and prevention. |
|
|
Beksac MS, Tanacan A, Ozten G, Cakar AN (2020). Low-dose low-molecular-weight heparin prophylaxis against obstetrical complications in pregnancies with metabolic and immunological disorder-associated placental inflammation. The Journal of Maternal-Fetal and Neonatal Medicine pp. 1-8. |
|
|
Bernal AL (2003). Mechanisms of labour - biochemical aspects. BJOG: An International Journal of Obstetrics and Gynaecology 110(4):39-45. |
|
|
Bezold KY, Karjalainen MK, Hallman M, Teramo K, Muglia LJ (2013). The genomics of preterm birth?: from animal models to human studies. Genome Medicine 5(4):1-11. |
|
|
Blencowe H, Cousens S, Chou D, Oestergaard M, Say L, Moller A, Kinney M (2013). Born Too Soon?: The global epidemiology of 15 million preterm births 10(1):1-14. |
|
|
Boyle AK, Rinaldi SF, Norman JE, Stock SJ (2017). Preterm birth: Inflammation, fetal injury and treatment strategies. Journal of Reproductive Immunology 119:62-66. |
|
|
Bulletti C, De Ziegler D (2005). Uterine contractility and embryo implantation. Current Opinion in Obstetrics and Gynecology 17(3): 265-276. |
|
|
Bulletti C, De Ziegler D, Polli V, Diotallevi L, Del Ferro E, Flamigni C. (2000). Uterine contractility during the menstrual cycle. Human Reproduction 15(1):81-89. |
|
|
Burguet A., Kaminski, M, Abraham-Lerat L, Schaal JP, Cambonie G, Fresson J, Grandjean H, Truffert P, Marpeau L, Voyer M, Rozé JC, Treisser A, Larroque B (2004). The complex relationship between smoking in pregnancy and very preterm delivery. Results of the Epipage study. BJOG: An International Journal of Obstetrics and Gynaecology 111(3):258-265. |
|
|
Challis JRG, Sloboda DM, Alfaidy N, Lye SJ, Gibb W, Patel FA, Whittle WL, Newnham JP (2002). Prostaglandins and mechanisms of preterm birth. Reproduction-Cambridge 124(1):1-17. |
|
|
Chatterjee J, Gullam J, Vatish M, Thornton S (2007). The management of preterm labour. Archives of Disease in Childhood-Fetal and Neonatal Edition 92(2):F88-F93. |
|
|
Cogswell ME, Weisberg P, Spong C (2003). Cigarette smoking, alcohol use and adverse pregnancy outcomes: Implications for micronutrient supplementation. Journal of Nutrition 133(5):1722-1731. |
|
|
Dickinson JE, Hart RJ, Pennell CE, Arrese CA, Keelan JA (2014). Strategies to prevent preterm birth. Frontiers in Immunology 5(11):1-12. |
|
|
Driscoll DNO, Mcgovern M, Greene CM, Molloy EJ(2018). Gender disparities in preterm neonatal outcomes. Acta Paediatrica pp. 1494-1499. |
|
|
Driul L, Londero AP, Vogrig E, Bertozzi S, Fachechi G, Forzano L (2014). Therapy side-eff ects and predictive factors for preterm delivery in patients undergoing tocolysis with atosiban or ritodrine for threatened preterm labour. Journal of Obstetrics and Gynaecology 34(8):684-689. |
|
|
Duckitt K, Thornton S, O'Donovan OP, Dowswell T (2014). Nitric oxide donors for treating preterm labour. Cochrane Database of Systematic Reviews (5). |
|
|
Dunk CE, Gellhaus A, Drewlo S, Baczyk D, Po AJG, Kingdom JCP, Lye SJ (2012). The Molecular Role of Connexin 43 in Human Trophoblast Cell Fusion Biology of Reproduction 86(4):115-1. |
|
|
Fanchin R, Ayoubi JM (2009). Uterine dynamics: impact on the human reproduction process. Reproductive biomedicine online 18:S57-S62. |
|
|
Farina L, Winkelman C (2005). A review of the role of proinflammatory cytokines in labor and noninfectious preterm labor. Biological Research for Nursing 6(3):230-238. |
|
|
Felice P, Alberto I, John RGC (2010). Neuroendocrine Mechanisms in Pregnancy and Parturition. Endocrine Reviews 31(6):783-816. |
|
|
Fifer, WP, Fingers ST, Youngman M, Gomez-Gribben E, Myers MM (2009). Effects of alcohol and smoking during pregnancy on infant autonomic control. Developmental Psychobiology 51(3):234-242. |
|
|
Galal M, Symonds I, Murray H, Petraglia F, Smith R (2012). Postterm pregnancy. Facts, Views and Vision in ObGyn 4(3):175-187. |
|
|
Galindo-Sevilla N, Reyes-Arroyo F, Mancilla-Ramírez J (2019). The role of complement in preterm birth and prematurity. Journal of Perinatal Medicine 47(8):793-803. |
|
|
Gestel IVan, Ijland MM, Hoogland HJ, Evers JLH (2003). Endometrial wave-like activity in the non-pregnant uterus. Human reproduction update 9(2):131-138. |
|
|
Goldenberg RL, Culhane JF, Iams JD, Romero R(2009). Preterm Birth 1 Epidemiology and causes of preterm birth. Obstetric Anesthesia Digest 29(1):6-7. |
|
|
Green ES, Arck PC (2020). Pathogenesis of preterm birth: bidirectional inflammation in mother and fetus. Seminars in Immunopathology 42(4):413-429. |
|
|
Haas DM, Benjamin T, Sawyer R, Quinney SK (2014). Short-term tocolytics for preterm delivery - Current perspectives. International Journal of Women's Health 6(1):343-349. |
|
|
Hamilton SA, Tower CL, Jones RL (2013). Identification of Chemokines Associated with the Recruitment of Decidual Leukocytes in Human Labour: Potential Novel Targets for Preterm Labour 8(2):e56946. |
|
|
Haram K, Helge J, Mortensen S, Morrison JC (2014). Tocolysis for acute preterm labor?: does anything work. The Journal of Maternal-Fetal & Neonatal Medicine 28(4):371-378.. |
|
|
Hubinont C, Debieve F (2011). Prevention of preterm labour: 2011 update on tocolysis. Journal of Pregnancy P 941057. |
|
|
Hutchinson JL, Rajagopal SP, Yuan M, Norman JE (2014). Lipopolysaccharide promotes contraction of uterine myocytes via activation of Rho / ROCK signaling pathways. The FASEB Journal 28(1):94-105. |
|
|
Ibrahim M, Ramy A, Ramy M, Younis MA (2010). Progesterone supplementation for prevention of preterm labor?: A randomized controlled trial. Middle East Fertility Society Journal 15(1):39-41. |
|
|
Ilicic M, Zakar T, Paul JW, Paul JW (2020). The Regulation of Uterine Function During Parturition: an Update and Recent Advances. Reproductive Sciences 27(1):3-28. |
|
|
Illamola SM, Amaeze OU, Krepkova LV, Birnbaum AK, Karanam A, Job KM, Bortnikova VV, Sherwin CMT, Enioutina EY (2019). Use of herbal medicine by pregnant women: What physicians need to know. Frontiers in Pharmacology 10:1483. |
|
|
Challis JR, Lockwood CJ, Myatt L, Norman JE, Strauss JF, Petraglia F (2009). Inflammation and pregnancy. Reproductive sciences, 16(2):206-215. |
|
|
Kashanian M, Akbarian AR, Soltanzadeh M (2005). Atosiban and nifedipin for the treatment of preterm labor. nternational Journal of Gynecology and Obstetrics 91(1):10-14. |
|
|
Kashanian M, Bahasadri S, Zolali B (2011). International Journal of Gynecology and Obstetrics Comparison of the ef fi cacy and adverse effects of nifedipine and indomethacin for the treatment of preterm labor. International Journal of Gynecology and Obstetrics 113(3):192-195. |
|
|
Kent AL, Children G, Wright IM, Abdel-latif ME (2011). Mortality and adverse neurologic outcomes are greater in preterm male infants. Pediatrics 129(1):124-131. |
|
|
Kim CJ, Romero R, Chaemsaithong P, Kim JS (2016). Chronic Inflammation of the Placenta: Definition, Classification, Pathogenesis, and Clinical Significance. HHS Public Access American Journal of Obstetrics and Gynecology 213(313):1-42. |
|
|
Koob TJ, Lim JJ, Massee M, Zabek N, Denozie G. (2014). Clinical Device Related Article Properties of dehydrated human amnion / chorion composite grafts?: Implications for wound repair and soft tissue regeneration pp. 1353-1362. |
|
|
Kosinski P, Luterek K, Lipa M, Wielgos M (2019). The use of atosiban prolongs pregnancy in patients treated with fetoscopic endotracheal occlusion ( FETO). Journal of perinatal medicine 47(9):910-914. |
|
|
Kota SK, Gayatri K, Jammula S, Kota SK, Krishna SV, Meher LK, Modi KD (2013). Endocrinology of parturition. Indian Journal of Endocrinology and Metabolism 17(1):50-59. |
|
|
Kronemyer B (2019). Preterm birth phenotypes in women with autoimmune disease. Contemporary OB/GYN 64(11):19-19. |
|
|
Larsen B, Hwang J (2011). Progesterone interactions with the cervix: translational implications for term and preterm birth. Infectious diseases in obstetrics and gynecology 2011. |
|
|
Lotfabadi LH (2013). The role of vitamin C in prevention of preterm premature rupture of membranes. Iranian Red Crescent Medical Journal 15(2):113-116. |
|
|
Loudon JA, Groom KM, Bennett PR (2003). Prostaglandin inhibitors in preterm labour. Best Practice & Research Clinical Obstetrics & Gynaecology 17(5):731-744. |
|
|
Ma M, Zhu M, Zhuo B, Li L, Chen H, Xu L, Wu Z, Cheng F, Xu L, Yan J (2020). Use of complete blood count for predicting preterm birth in asymptomatic pregnant women?: A propensity score-matched analysis. Journal of clinical laboratory analysis 34(8):e23313. |
|
|
Malhotra C, Jain AK (2014). Human amniotic membrane transplantation: Different modalities of its use in ophthalmology. World Journal of Transplantation 4(2):111. |
|
|
Mandruzzato G, Alfirevic Z, Gruenebaum A, Heimstad R, Heinonen S, Levene M, Salvesen K, Saugstad O, Skupski D (2010). Guidelines for the Management of Postterm Pregnancy 38:111-119. |
|
|
Mccormick MC, Litt JS, Smith VC, Zupancic JAF (2011). Prematurity?: An Overview and Public Health Implications. Annual Review of Public Health 33:367-379. |
|
|
Mol BWJ, Pajkrt E, Oudijk MA (2016). Prevention of preterm delivery?: current challenges and future prospects. International Journal of Women's Health 8:633. |
|
|
Nadeem L, Shynlova O, Matysiak-Zablocki E, Mesiano S, Dong X, Lye S (2016). Molecular evidence of functional progesterone withdrawal in human myometrium. Nature communications 7(1):1-9. |
|
|
Neil JJ, Volpe JJ (2018). Encephalopathy of prematurity: Clinical-neurological features, diagnosis, imaging, prognosis, therapy. In Volpe's Neurology of the Newborn (pp. 425-457). Elsevier. |
|
|
Newnham JP (2017). Recent advances in the prevention of preterm birth [ version 1?; referees?: 2 approved ] Jeff A Keelan Referee Status 6(0):1-11. |
|
|
Niebyl JR, Witter FR. (1986). Neonatal outcome after indomethacin treatment for preterm labor. American Journal of Obstetrics and Gynecology 155(4):747-749. |
|
|
Noakes PS, Michaelis LJ (2013). Innate and adaptive immunity. In Diet, Immunity and Inflammation. In Woodhead Publishing. Woodhead Publishing Limited. |
|
|
Norman JE. (2020). Progesterone and preterm birth. International Journal of Gynecology and Obstetrics 150(1):24-30. |
|
|
Norwitz ER, Caughey AB (2011). Progesterone Supplementation and the Prevention of Preterm Birth 4(2):60-72. |
|
|
Olson DM, Ammann C (2015). Role of the prostaglandins in labour and prostaglandin receptor inhibitors in the prevention of preterm labour. Front Biosci 12(1):1329-1343. |
|
|
Osman I, Young A, Ledingham MA, Thomson AJ, Jordan F, Greer IA, Norman JE (2003). Leukocyte density and pro-inflammatory cytokine expression in human fetal membranes, decidua, cervix and myometrium before and during labour at term. Molecular Human Reproduction 9(1):41-45. |
|
|
Ousey JC, Fowden AL (2012). Prostaglandins and the regulation of parturition in mares. Equine Veterinary Journal (44):140-148. |
|
|
Perales A (2018). Electrohysterography in the diagnosis of preterm birth?: a review. Physiological Measurement 39(2):02TR01. |
|
|
Phillips RJ, Fortier MA, Bernal AL (2014). Prostaglandin pathway gene expression in human placenta, amnion and choriodecidua is differentially affected by preterm and term labour and by uterine inflammation. BMC Pregnancy and Childbirth 14(1):1-14. |
|
|
Phung, J., Paul, J., & Smith, R. (2020). Maintenance of pregnancy and parturition. In Maternal-Fetal and Neonatal Endocrinology pp. 169-187. Academic Press. |
|
|
Pierce SL (2010). The role and regulation of small conductance CA2 + activated K + channel subtype 3 in myometrial contraction and placental development. |
|
|
Purisch SE, Gyam C (2017). Seminars in Perinatology Epidemiology of preterm birth. 41:387-391. |
|
|
Rabotti C, Sammali F, Kuijsters N, Schoot B, Kortenhorst M, Mischi M (2015). Analysis of uterine activity in nonpregnant women by electrohysterography: a feasibility study. pp. 5916-5919. |
|
|
Ramos BR de A, Mendes ND, Tanikawa AA, Amador MAT, Santos NP C dos, Santos SEB dos, Castelli EC, Witkin SS, Silva MG da (2016). Ancestry informative markers and selected single nucleotide polymorphisms in immunoregulatory genes on preterm labor and preterm premature rupture of membranes: A case control study. BMC Pregnancy and Childbirth 16(1):1-11. |
|
|
Visse R, Nagase H (2003). Matrix Metalloproteinases and Tissue Inhibitors of Metalloproteinases. Circulation Research 92(8):827-839. |
|
|
Rubens CE, Sadovsky Y, Muglia L, Gravett MG, Lackritz E, Gravett C. (2014). STATE OF THE ART REVIEW Prevention of preterm birth: Harnessing science to address the global epidemic. Science Translational Medicine 6(262):262sr5-262sr5. |
|
|
Sakai M, Tanebe K, Sasaki Y, Momma K, Yoneda S, Saito S (2001). Evaluation of the tocolytic effect of a selective cyclooxygenase-2 inhibitor in a mouse model of lipopolysaccharide-Induced Preterm Delivery 7(6):595-602. |
|
|
Sammali F, Pertronella N, Kuijsters M, Schoot BC, Mischi M, Rabotti C (2018). Feasibility of Transabdominal Electrohysterography for Analysis of Uterine Activity in Nonpregnant Women. |
|
|
Sanborn BM, Ku CY, Shlykov S, Babich L(2005). Molecular signaling through G-protein-coupled receptors and the control of intracellular calcium in myometrium. Journal of the Society for Gynecologic Investigation: 12(7):479-487. |
|
|
Sanborn BM (2001). Uterine contractility symposium?: The Litchfield Lecture Hormones and calcium?: mechanisms controlling uterine smooth muscle contractile activity. Experimental physiology 86(2):223-237. |
|
|
Sanu O, Lamont RF (2010). Critical appraisal and clinical utility of atosiban in the management of preterm labor. Therapeutics and Clinical Risk Management 6(191):191-199. |
|
|
Schleussner E, Lehmann T, Kähler C, Schneider U (2014). Impact of the nitric oxide-donor pentaerythrityl- tetranitrate on perinatal outcome in risk pregnancies?. Journal of Perinatal Medicine 42(4):507-514. |
|
|
Schwarz MK, Page P (2003). Preterm Labour?: An Overview of Current and Emerging Therapeutics. Current Medicinal Chemistry 10(15):1441-1468. |
|
|
Shynlova O, Lee Y, Srikhajon K, Lye SJ (2013). Physiologic Uterine Inflammation and Labor Onset?: Integration of Endocrine and Mechanical Signals 20(2):154-167. |
|
|
Silva E, Leitão S, Henriques S, Kowalewski MP, Hoffmann B, Ferreira- Dias G, Lopes da Costa L, Mateus L (2010). Gene transcription of TLR2, TLR4, LPS ligands and prostaglandin synthesis enzymes are up-regulated in canine uteri with cystic endometrial hyperplasia-pyometra complex. Journal of Reproductive Immunology 84(1):66-74. |
|
|
Souza RT, Cecatti JG, Passini R, Tedesco RP, Lajos GJ, Nomura ML, Rehder PM., Dias TZ, Haddad SM, Pacagnella RC, Costa ML (2016). The burden of provider-initiated preterm birth and associated factors: Evidence from the Brazilian Multicenter study on preterm birth (EMIP). PLoS ONE 11(2):1-20. |
|
|
Soon B T (2012). The global action report on preterm birth. Geneva: World Health Organization. |
|
|
Stabile I, Chard T, Grudzinskas G (2000). Premature Labour. In Clinical Obstetrics and Gynaecology pp. 59-61. Springer London. |
|
|
Stelzl P, Kehl S, Rath W (2019). Maintenance tocolysis?: a reappraisal of clinical evidence. Archives of Gynecology and Obstetrics 0123456789. |
|
|
Tanacan A, Beksac MS, Orgul G, Duru S, Sener B, Karaagaoglu E (2019). Impact of extractable nuclear antigen, anti-double stranded DNA, antiphospholipid antibody, and anticardiolipin antibody positivity on obstetrical complications and pregnancy outcomes. Human antibodies 27(2):135-141. |
|
|
Tanacan E (2021). A cutoff value for the Systemic Immune-Inflammation Index in determining activity of Behc ß et disease pp. 286-291. |
|
|
Terzidou V (2007). Biochemical and endocrinological preparation for parturition. Best Practice and Research: Clinical Obstetrics and Gynaecology 21(5):729-756. |
|
|
Thoene M, Van Ormer M, Yuil-Valdes A, Bruett T, Natarajan SK, Mukherjee M, Thompson M, Nordgren TM, Van Lippevelde W, Overby NC, Adu-Bonsaffoh K, Anderson-Berry A, Hanson C (2020). Fat-soluble nutrients and Omega-3 fatty acids as modifiable factors influencing preterm birth risk. Placenta 98:38-42 |
|
|
Vadillo-ortega F, Estrada-gutie G (2005). Role of matrix metalloproteinases in preterm labour. BJOG: An International Journal of Obstetrics and Gynaecology 112:19-22. |
|
|
Vannuccini S, Bocchi C, Severi FM, Challis JR, Petraglia F (2016). Endocrinology of human parturition. Annales d'Endocrinologie 77(2):105-113. |
|
|
Vinturache AE, Gyamfi-Bannerman C, Hwang J, Mysorekar IU, Jacobsson B, Collaborative TPBI (2016). Maternal microbiome-a pathway to preterm birth. In Seminars in Fetal and Neonatal Medicine 21(2):94-99). WB Saunders. |
|
|
Vogel JP, Nardin JM, Dowswell T, West HM, Oladapo OT (2014). Combination of tocolytic agents for inhibiting preterm labour. Cochrane Database of Systematic Reviews (7). |
|
|
Walker SK, Hartwich KM, Robinson JS (2000). Long-term effects on offspring of exposure of oocytes and embryos to chemical and physical agents. Human Reproduction Update 6(6):564-577. |
|
|
Wang H, Hu YF, Hao JH, Chen YH, Su PY, Wang Y, Yu Z, Fu L, Xu Y Y, Zhang C, Tao FB, Xu, DX (2015). Maternal zinc deficiency during pregnancy elevates the risks of fetal growth restriction: A population-based birth cohort study. Scientific Reports 5(7):1-10. |
|
|
Wei SQ, Fraser W, Luo ZC (2010). Inflammatory cytokines and spontaneous preterm birth in asymptomatic women: A systematic review. Obstetrics and Gynecology 116(2):393-401. |
|
|
Whitaker E, Murray S, Norman JE (2019). Clinical Interventions for the Prevention and Management of Spontaneous Preterm Birth in the Singleton Fetus pp. 311-324. |
|
|
Wise CA, Sharma S (2010). Current understanding of genetic factors in idiopathic scoliosis. The Genetics and Development of Scoliosis 112(3):167-190. |
|
|
Yang F, Zheng Q, Jin L (2019). Dynamic Function and Composition Changes of Immune Cells During Normal and Pathological Pregnancy at the Maternal-Fetal Interface 10(10):1-15. |
|
|
You X, Xu C, Lu J, Zhu X, Gao L, Cui X, Li Y, Gu H (2011). Expression of Cystathionine b -synthase and Cystathionine c -lyase in Human Pregnant Myometrium and Their Roles in the Control of Uterine Contractility 6(8):1-9. |
|
|
Zhang Q, Ananth CV, Li Z, Smulian JC (2009). Maternal anaemia and preterm birth: A prospective cohort study. International Journal of Epidemiology 38(5):1380-1389. |
|
|
Ziegler DDE, Bulletti C, Fanchin R, Epiney M (2001). Contractility of the Nonpregnant Uterus. Annals of the New York Academic of Sciences 943(1):172-184. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


