Full Length Research Paper

ABSTRACT
Coffee supports livelihoods of approximately 125 million families worldwide and over 700,000 households in Kenya. The epidemics of Coffee berry disease (CBD), caused by Colletotrichum kahawae, destroy up to 80% of the developing berries on susceptible varieties. The control of the disease using chemicals accounts for 30 to 40% of the production cost and contributes to environment pollution, hence the use of resistant varieties. Resistance to CBD is conferred by three genes; R, T that are dominant and k which is recessive, from coffee varieties Rume Sudan (RS), Hibrido de Timor (HDT) and K7 respectively. Although the T gene has been mapped, there is need for genetic mapping of the other genes to improve selection efficiency. The objective of this study was to evaluate F2 populations of RS x SL28 for their suitability to genetic mapping of the R gene in RS. Resistance to CBD was evaluated by hypocotyl inoculation on their F3 progenies. The data was subjected to Analysis of Variance (ANOVA) and Chi Square (χ²) test. The ANOVA result showed significant differences (P≤0.05) between the genotypes to CBD resistance. The phenotypic ratio of resistance to susceptible plants fitted the 3:1 monohybrid inheritance ratio for a dominant gene using the χ² test (χ² = 1.0565 and P=0.30207, P≤0.05), hence confirming the suitability of the F2 populations for the identification of the DNA marker for R gene in RS.
Key words: Rume Sudan, SL 28, Coffee Berry Disease, mapping population, hypocotyl, inoculation.
INTRODUCTION
Coffee (Coffea spp.) is among the most important commodities in the tropical countries of the world (Vieira et al., 2019). It is commercially grown in more than 10.5 million ha in 80 different countries worldwide (Van der Vossen et al., 2015). It supports livelihoods of approximately 125 million families in coffee producing countries (Zhou et al., 2016). Coffee is an important export crop and a major foreign exchange earner for Kenya, supporting over 700,000 smallholder farmers that are organized into about 435 farmers’ co-operative societies. The cooperative societies account for 80% of the total coffee production while the remaining 20% is produced by 3000 coffee plantations (Kathurima, 2013; Gimase, 2014). Coffee ranks fifth in terms of economic importance after tea, tourism, horticultural sub-sector and diaspora remittance. It contributes about 1% to GDP and 8% of the total agricultural export revenue (Kenya National Bureau of Statistics, 2019). Of the 70% of Kenya’s workforce engaged in agriculture, 30% are employed by the coffee industry (Minai et al., 2014). Kenya is known for production of some of the worlds’ top grade and highly valued Arabica coffee beans that are usually used in small quantities by international coffee roasters to moderate and improve their blends (Kathurima, 2013).
Coffea arabica L., is a tetraploid (2n = 4x = 44) and 95% self-fertile (Bertrand et al., 2003). Coffee was introduced in Kenya at the beginning of 20th Century by missionaries (Omondi et al., 2016). It is mainly grown in three regions, the East of Rift Valley (areas around Mount Kenya, the Aberdare ranges and Machakos), West of Rift Valley (Kisii highlands, area around Mt Elgon and the North of the Rift valley) and Taita Hills in the coast (Kathurima, 2013).
Coffee berry disease caused by the fungal pathogen Colletotrichum kahawae, is a specialized hemibiotrophic pathogen of C. arabica L. (Vieira et al., 2019). Unlike other Arabica coffee diseases, CBD is still restricted to the African continent despite favourable climatic conditions in certain high-altitude Arabica coffee growing areas of Latin America and Asia (Agwanda et al., 1997; Van der Vossen et al., 2015). Colletotrichum kahawae infects green berries at the rapid expansion stage (4-16 weeks after flowering) and may also attack mature berries, 28 weeks after flowering (Gichimu et al., 2014, CRI, 2016). Epidemics of this disease can quickly destroy 50–80% of the developing berries on susceptible Arabica coffee cultivars during prolonged wet and cool weather conditions (Hindorf and Omondi, 2011). Preventive control by frequent fungicide sprays account for 30–40% of total production costs and leads to environmental pollution (Gichuru et al., 2008). Crop loss and cost of CBD control in Africa is estimated to be 300 – 500 million USD (Van der Vosen and Walyaro, 2009).
The first case of CBD in Kenya was reported in 1922, in newly established coffee plantations on the slope of Mt Elgon in Western Kenya (McDonald, 1926). The disease then spread to other parts of the country, East of Rift Valley by 1939 and to all other main coffee growing zones by 1951. It was from Kenya that CBD spread to Angola in 1930, Zaire in 1937, Cameroon between 1955 and 1957, Uganda in 1959, Tanzania in 1964, Ethiopia in 1971 and Malawi in 1985 (Hindorf and Omondi, 2011). Breeding for resistance to CBD in Kenya, started in 1971, following a serious outbreak of CBD in 1967-68 (Van Der Vossen and Walyaro, 1981). The CBD epidemics experienced at that time affected all the Kenyan commercial varieties and threatened to wipe out the coffee industry in the country (Van der Vosen and Walyaro, 1981; Hindorf and Omondi, 2011).
Marker-assisted selection is one of the best approaches to reduce the period taken to develop coffee varieties (Moncada et al. 2016). To implement this technique, one requires to develop a genetic map for markers that are associated with traits of interest, a process that also involves the development of a mapping population (Baison, 2014). The simplest population is the F2 genotypes (Schneider, 2005). The mode of reproduction influences the choice of mapping populations and the relative ease of raising such populations. An ideal mapping population therefore, should be derived from parents with a wide variation in the trait to be analyzed (Gichuru, 2007). Self-fertile naturally inbreeding plants of Arabica coffee attain a high degree of homozygosity and well-varied pure line parents for generating mapping populations (Gichuru, 2007).
The discovery of hypocotyl infection on a six-week-old seedling using artificial inoculation with Colletotrichum kahawae spores contributes significantly to Arabica coffee breeding by shortening the time required to identify resistant progenies from crosses involving resistant and susceptible donors (Van Der Vossen et al., 1976; Agwanda et al., 1997; Gichuru et al., 2008). The method was found to be a reliable pre-selection test whose result was significantly correlated (r2=0.73-80, P≤0.05) with mature plant resistance in the field (Van Der Vossen et al., 1976).
Inheritance studies on Arabica coffee genotypes have identified three genes that confer resistance to CBD in C. arabica as R-gene in the variety Rume Sudan, T-gene in Hibrido de Timor and k-gene in K7 (Van der Vossen and Walyaro, 1980). The T gene has an intermediate gene action, with the R gene dominant and the k gene recessive; therefore, the k gene only confers partial resistance to CBD (Van der Vossen and Walyaro, 2009).
Genetic resistance to CBD has been characterized by various studies in Kenya. The T gene was mapped by Agwanda et al. (1997) using random amplified polymorphic DNA (RAPD) markers while Gichuru et al. (2008) mapped the first locus for resistance to Colletotrichum kahawae using simple sequence repeats (SSR) markers and amplified fragment length polymorphisms (AFLP) markers and christened it as Ck-1. The locus was found to be linked to the highly repetitive and informative SSR primer locus (Sat 235) that has been widely adopted for marker assisted selection (MAS) in Arabica coffee (Gichimu et al. 2014; Mtenga, 2016; Alkimim et al. 2017). Although molecular markers for the T-gene (Ck-1) gene in HDT was detected, similar molecular research studies remain necessary for the detection and mapping of R genes in Rume Sudan. This will increase selection efficiency in the rapid development of CBD resistant varieties to CBD that meet consumers preference for Arabica coffee growing countries in Africa and breeding programmes in Latin America, where CBD has not been reported but there remains a possible likelihood of its occurrence (Van der Vossen et al., 2015).
Evaluation of F2 populations to determine their suitability for mapping of resistance to diseases in coffee has been carried out by various studies. Gichuru (2007) evaluated two F2 populations derived from CBD resistance donor parent Catimor and susceptible cultivar SL 28; Brito et al. (2010) F2 populations from HDT UFV 427-15 and the susceptible cultivar Catuai Amarelo UFV 2143-236 for their segregation on resistance to race II Heimillea vastatrix, the causal agent of coffee leaf rust. Similarly, Diola et al. (2011, 2013) studied an F2 population obtained by crossing HDT UFV 427-15 (resistant) with Catuai Amarelo IAC 30 (susceptible) segregating for a dominant gene that confers resistance on coffee to race II of Heimillea vastatrix while Pestana et al. (2015) examined F2 population of a cross between Catuai Amarelo IAC 64 (UFV 2148-57), a susceptible Arabica coffee variety and HDT UFV 443-03 as a donor variety of resistance to Heimillea vastatrix.
Breeding programs in various crops have led to release of genotypes with improved traits that often breaks up after a short period as different genes for resistance acts against different isolates, races or biotypes (Mekonnen et al., 2017). The process of accumulating various genes to a given genotype (pyramiding) broadens the number of races or isolates that one variety can resist or tolerate at the same time (Sundaram et al., 2009). Pyramiding of genes for resistance using conventional screening methods is limited by the dominance effects of genes governing disease resistance (Arunakumari et al., 2016); for DNA markers, it is possible to accurately identify genes of interest of the progenies at each generation, thus making pyramiding process faster and more efficient (Zhao et al., 2014). Despite the fact that resistance to CBD is controlled by three genes (Van der Vossen and Walyaro, 1980), only the T gene has been mapped (Gichuru et al., 2008) and adopted for MAS (Alkimim et al. 2017). It is therefore necessary to identify DNA markers for R and k genes to increase selection efficiency in the rapid development of CBD resistant varieties (Van der Vossen et al., 2015). The objective of this study was to evaluate F2 populations derived from crosses between C. arabica varieties Rume Sudan and SL 28 for resistance to CBD and determine their suitability of genetic mapping of R gene that confer resistance to CBD in C. arabica L.
MATERIALS AND METHODS
This study was conducted at Coffee Research Institute (CRI) of the Kenya Agricultural and Livestock Research Organization (KALRO), Ruiru Centre. Ruiru is located within the upper midland (UM2) at 1° 06'S and 36° 45'E and at an altitude of 1620 m above sea level. The rainfall pattern is bimodal with 1063 mm per annum and the annual average temperature is 19°C with a range of 12.8 to 25.2°C (Jaetzold et al., 2006).
Study materials
This study utilized a total of 108 genotypes comprising 106 F2 genotypes and their parents, Rume Sudan and SL28. F2 segregating population of a cross between C. arabica varieties Rume Sudan (RS) and SL 28 was developed and established at CRI. In the development of RS x SL28 F2 populations, the cultivar RS was used as female parent. Rume Sudan is a normal Arabica variety that is believed to possess the R gene in the R locus that confers resistance to CBD while the male parent SL 28 is C. arabica cultivar highly susceptible to CBD (Hindorf and Omondi, 2011).
Evaluation for resistance to CBD and classification of F2 populations
The 108 genotypes were classified based on resistance to CBD of their F3 progenies using hypocotyl inoculation method on a scale of 1 – 12, as described by Van der Vossen et al. (1976) (Table 1). The F2 plants were selfed to generate F3 progenies for two seasons (2017 and 2018). For each season, ripe and healthy F3 berries were harvested from each of the F2 Rume Sudan x SL 28 individual genotype. The harvested berries were manually pulped by hand-squeezing them between the fingers. The seeds were fermented for about 16 h, washed, dried to a moisture content of 15% and then the parchment was removed by hands. Three hundred seeds were planted in sterilized sand in plastic boxes and kept at room temperature under laboratory conditions. Twenty seedlings of the susceptible SL 28 were also sown alongside the test seedlings in each box. Coffee variety SL 28 was used as a susceptible control to verify success of infection. The experiment was set out in the laboratory in a completely randomized design (CRD) with three replicates each of 100 seeds. Watering was carried out twice per week using distilled water to ensure that the sand remained moist but not water logged. After 6 weeks, the germinated hypocotyl seedlings with unopened cotyledons were uprooted and immediately replanted in similar but clean boxes filled with sterilized sand at a spacing of 2.5 cm x 2.5 cm.

Isolates of Colletotrichum kahawae were obtained from freshly infected coffee berries in the field and multiplied on malt extract agar (MEA) for a period of about 15 days in the laboratory. To stimulate conidia production, isolates were cultured on coffee leaf extract agar medium for 7 days under a photoperiod of 12 h at 22°C and then sub-cultured on 90-mm polystyrene petri dishes containing malt extract agar (40 g L-1, MEA; Oxoid) for 7 days under the same photoperiod (Vieira et al., 2019). Inoculum was obtained by dislodging and harvesting the conidia by flooding the plate with 5 mL of sterile distilled water and the suspensions passed through four layers of sterile muslin cloth to remove mycelia (Vieira et al., 2019). Concentrations of spore suspensions were determined using a haemocytometer (NEUBAUER Scientific International, Germany).
Six weeks after sowing the F3 seeds in the sand boxes, inoculum suspension was adjusted and standardized to a concentration of 2 x 106 conidia per ml (Van der Vossen et al., 1976; Viera et al., 2019). The hypocotyls were inoculated using a hand sprayer, spraying them twice at 48-h interval with the inoculum. After every spray interval, the seedlings were incubated in the dark by covering them with black polythene sheet for 48 h at room temperature and then transferred to a temperature-controlled room at 18 to 20°C for 2 weeks. The seedlings were transferred back to room temperature for one week, after which disease symptom severity rating were carried out. The incubation period was determined by the full expression of disease on SL 28. Each seedling was assessed based on expression of disease symptoms on the hypocotyls. Average infection (AI) per replicate was calculated as follows:
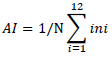
where, i is the disease class, ni is the number of seedlings in class i, N is the total number of seedlings scored (Van der Vossen et al., 1976) and mean grade data computed for each genotype.
Chi squared tests for goodness of fit on Mendelian monohybrid inheritance ratio for phenotypic segregation of 3:1 (resistant to susceptible) for a dominant gene was used to confirm genetic hypothesis on the mode of inheritance for CBD resistance of the genotypes (Fazel-Najafabadi et al., 2015; Kim and Reinke, 2019).
RESULTS
Phenotypic segregation of the F2 Population for CBD resistance
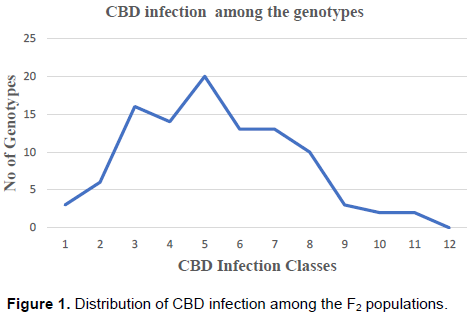
The SL 28 seedlings were ranked highly susceptible to CBD with a rating between 11 and 12 and a mean of 11.9 (Table 2), an indication that the infection was highly successful. Rume Sudan recorded a disease rating of 2.97 and was considered highly resistant to CBD. The F2 genotypes segregated showing various levels of resistance and susceptibility, however, no genotypes had a score of 12 (Table 2). The mean infection score for all the genotypes was 5.75. Based on the hypocotyl inoculation results, the genotypes were classified into two phenotypic classes by comparing the infection rates of the F2 populations with SL 28. Seedlings with ratings of 7 and 12 were considered susceptible and those rated from 1 – 6, considered resistant (Van Der Vossen et al., 1976). The frequency curve of CBD resistance on the F2 genotypes showed a continuous distribution from grade 1 to 12, but was slightly skewed towards more resistance genotypes (Figure 1).
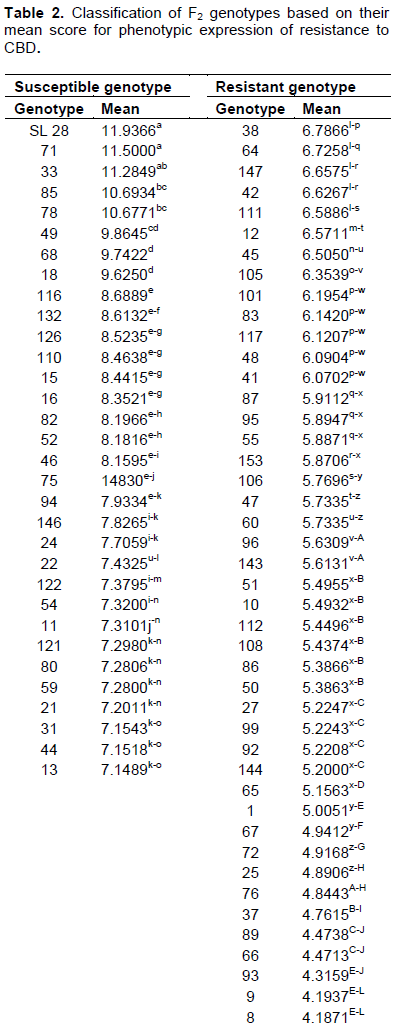
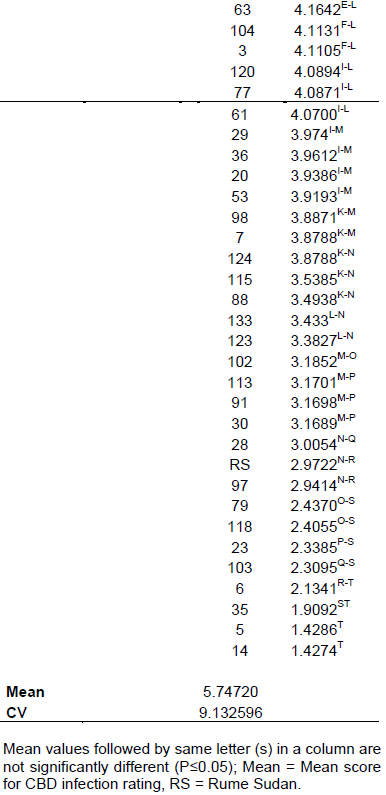
The mean data on phenotypic segregation for CBD resistance of RS x SL28 F2 populations was subjected to analysis of variance using SAS statistical software (Version 9) and means separated using least significant difference (LSD). The result showed significant variation (P≤0.05) among the genotypes for resistance to CBD (Table 2). Three genotypes namely 35, 5 and 14 were significantly more resistant (P≤0.05) to CBD than Rume Sudan whereas two genotypes, 71 and 33 were not significantly different (P≤0.05) from the susceptible check (SL 28).
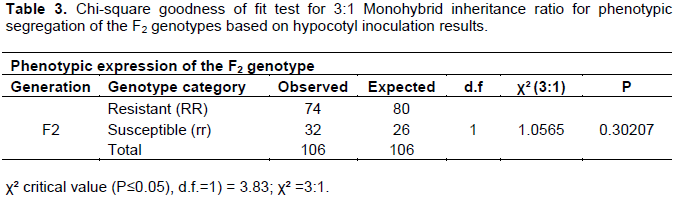
The mean data was subjected to Chi-square test in order to check for goodness-of-fit for the various Mendelian monohybrid inheritance ratios for the segregation of a dominant gene (Table 3). The segregation ratios of resistant to susceptible genes (R:S) is used by breeders to determine conformity of populations to the expected genetic segregations (Baison, 2014). The phenotypic segregation of CBD infection distribution fitted the 3:1 monohybrid ratio (χ² = 1.0565 and P=0.30207, P≤0.05) for the F2 mapping populations (Table 3).
DISCUSSION
The study revealed significant variations in the resistance to CBD within the F2 populations derived from RS x SL28 crosses. Related results were reported by Gichuru (2007) who observed significant variation in resistance to CBD among F2 population derived from CBD resistance donor parents Catimor and susceptible cultivar SL 28. Gichimu et al. (2014) observed variations in the resistance to CBD among coffee variety R11 parental genotypes and within different Ruiru 11 siblings. Mtenga (2016) also found significant differences for CBD resistance on progenies of a cross between Ethiopian accessions and C. arabica cultivar, KP423 for resistance to CBD.
The variety SL 28 had a high rating of 11 - 12 in the reaction to the inoculation of Colletotrichum kahawae, whereby at least most of the stem of the hypocotyl was affected, with more than one third of the stem found to have shriveled, leading to the death of the seedling and confirming its high susceptibility to the disease. A related result was reported by Gichimu et al. (2014) recorded a disease rate of 11.59 – 11.72 for SL 28; Gichuru (2007), who recorded 11.8 for the susceptible cultivar Catura; Omondi et al. (2001), 10.5 – 12 for SL 28 and Van Der Vossen et al. (1976), 10 - 12 for SL 28. A relatively recent study by Mtenga (2016), using cultivar KP423, which is a susceptible C. arabica commercial variety in Tanzania recorded the highest CBD scores among the F1 progenies. Rume Sudan was rated as highly resistant with a disease rating of 2.97. Similar results were reported by Van Der Vossen et al. (1976) with disease rating of 4.1 and Gichimu et al. (2014) who reported 4.6. Rume Sudan and SL 28 scores ranged from highly resistant class to most susceptible class rating, respectively. An ideal mapping population should be derived from parents with a large variation in the trait to be analyzed (Gichuru, 2007; Baison, 2014; Moncada et al. 2016). Therefore, in this study, Rume Sudan x SL 28 could be an ideal parental combination for a mapping population following their wide rating range for CBD resistance.
The phenotypic ratios of resistant to susceptible genotypes were 74:32 which fitted a 3:1 monohybrid inheritance ratio for a major/dominant gene (χ² = 1.0565 and P=0.30207, P≤0.05) for the F2 populations (Table 3). A study by Gichuru (2007) on a dominant T gene that confer resistance to CBD in HDT using two F2 population derived from susceptible cultivar SL 28 and Catimor as a donor of the resistance gene, revealed that the ratio of resistant to susceptible for two populations were 96:35 and 103:44 which fitted the 3:1 Mendelian ratio for a major gene action (χ² = 0.206; P=0.650 and χ² = 1.907; P=0.167) for the two respective populations. Brito et al. (2010) evaluated 160 F2 genotypes derived from a cross between the resistant genotype Hibrido de Timor UFV 427-15 and the susceptible cultivar Catuai Amarelo UFV 2143-236 for their segregation on resistance to race II Heimillea vastatrix, the causal agent for coffee leaf rust and reported 124:36 for R:S that fitted in the 3:1 Mendelian ratio for a dominant gene (χ²= 0.5336, P = 0.4652). Diola et al. (2011) reported phenotypic ratio of 166:58 that also fitted on the 3:1 segregation pattern expected for a single, dominant gene (χ²=0.09524) on F2 population from a crossing of Hibrido de Timor UFV 427-15 (resistant) with Catuaí Amarelo IAC 30 (susceptible), for a dominant gene that confers resistance on coffee to race II of Heimillea vastatrix. Diola et al. (2013) reported a Mendelian ratio of resistant to susceptible genotypes of 3:1, indicating that one gene is involved in the resistance of the HDT to pathotype of race II of Heimillea vastatrix using 224 F2 plants derived from resistant parent HDT UFV 427-15 and the susceptible parent Catuaí Amarelo UFV 2143-236 (IAC 30). In a related study, Pestana et al. (2015) also reported 3:1 (P≤0.05) ratio for dominant gene in an F2 population of a cross between Catuai Amarelo IAC 64 (UFV 2148-57), a susceptible Arabica coffee variety and Hibrido de Timor UFV 443-03, a donor variety for resistance Heimillea vastatrix. Kim and Reinke et al. (2019) reported expected phenotypic ratio of 3:1 (χ² = 4.15 and P = 0.12, P≤0.05) on F2 rice genotypes evaluated for resistance to bacterial blight.
The distribution of the infection rating among the F2 genotypes was close to normal with a skew toward the lower level of CBD infection. The skewness is a necessary for determination of Mendelian inheritance ratios. A study by Kim and Reinke et al. (2019) revealed a skewed result towards resistant genotypes on F2 rice genotypes evaluated for resistance to bacterial blight.
CONCLUSION
This study revealed significant variation in the symptoms of Colletotrichum kahawae among the F2 populations derived from Arabica coffee varieties Rume Sudan and SL 28. The CBD disease rating was skewed towards the resistant genotypes, demonstrating an ideal segregation of the populations for resistance. There was a significant contrast in terms of resistance to CBD between RS and SL 28, indicating that the choice of the parents was justified. This study also showed that the phenotypic segregation within the populations for resistant to susceptible genotypes fitted 3:1 Mendelian ratio expected from a dominant gene. The F2 populations derived from RS and SL 28 were suitable for genetic mapping of a dominant gene. Therefore, RS x SL 28 F2 populations are recommended for mapping of R gene in the variety RS that confers resistance to CBD to C. arabica L.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors thank the Director General, KALRO and the Institute Director, CRI who facilitated this study. The technical staffs of Breeding and Pathology section of CRI are also appreciated for their support in data collection. This work is published with permission of the Institute Director.
REFERENCES
|
Agwanda CO, Lashermes P, Trouslot P, Combes MC, Charrier A (1997). Identification of RAPD markers for resistance to coffee berry disease, Colletotrichum kahawae, in arabica coffee. Euphytica 97:241-248. |
|
|
Alkimim ER, Caixeta ET, Sousa TV, Pereira AV, Oliveira ACB, Zambolim L, Sakiyama NS (2017). Marker-assisted selection provides Arabica coffee with genes from other Coffea species targeting on multiple resistance to rust and coffee berry disease. MoleculaR Breeding 37(6):1-10. |
|
|
Arunakumari K, Durgarani CV, Satturu V, Sarikonda KR, Chittoor PDR, Vutukuri B, Laha GS, Nelli APK, Gattu S, Jamal M, Prasadbabu A, Hajira S, Sundaram RM (2016). Marker-Assisted Pyramiding of Genes Conferring Resistance against Bacterial Blight and Blast Diseases into Indian Rice Variety MTU1010. ScienceDirect Rice Science 23(6):306-316. |
|
|
Baison J (2014). Mapping and identification of disease resistance candidate genes in three Malus populations using SSRs, DArT and Infinium SNP markers and Illumina sequencing technology (PhD thesis), University of the Western Cape, South Africa. p. 343. |
|
|
Bertrand B, Guyot B, Anthony F, Lashermes P (2003). Impact of Coffea canephora gene introgression on beverage quality of C. arabica. Theoretical and Applied Genetics 107(3):387-394. |
|
|
Brito GG, Caixeta ET, Gallina AP, Zambolim EM, Zambolim L, Diola V, Loureiro ME (2010). Inheritance of coffee leaf rust resistance and identification of AFLP markers linked to the resistance gene. Euphytica 173:255-264. |
|
|
Coffee Research Institute (2016). Coffee Production Recommendation, 4th Ed. p 107. |
|
|
Diola V, Brito GG, Caixeta ET, Zambolim EM, Sakiyama NS, Loureiro ME (2011). High-density genetic mapping for coffee leaf rust resistance. Tree Genetics and Genomes 7(6): 1199-1208. |
|
|
Diola V, Brito GG, Caixeta ET, Pereira LFP, Loureiro ME (2013). A new set of differentially expressed signaling genes is early expressed in coffee leaf rust race II incompatible interaction. Functional and Integrative Genomics 13(3):379-389. |
|
|
Fazel-Najafabadi M, Peng J, Peairs FB, Simkova H, Kilian A, Lapitan NLV (2015). Genetic mapping of resistance to Diuraphis noxia (Kurdjumov) biotype 2 in wheat (Triticum aestivum L.) accession CI240. Euphytica 203:607-614. |
|
|
Gichimu BM, Gichuru EK, Mamati GE, Nyende AB (2014). Occurrence of Ck-1 gene conferring resistance to Coffee Berry Disease in Coffea arabica cv. Ruiru 11 and its parental genotypes. Journal of Agricultural and Crop Research 2(3): 51-61. |
|
|
Gichuru EK. (2007). Characterization of genetic resistance of Coffee berry disease (Colletotrichum kahawae, Waller and Bridge) in Arabica coffee (Coffea arabica L.) that is introgressed from Coffea canephora (PhD thesis), University of Nairobi, Kenya. P. 224. |
|
|
Gichuru EK, Agwanda CO, Combes MC, Mutitu EW, Ngugi ECK, Bertrand B, Lasherme, P (2008). Identification of molecular markers linked to a gene conferring resistance to Coffee berry disease (Colletotrichum kahawae) in Coffea arabica. Plant Pathology 57:1117-1124. |
|
|
Gimase JM (2014). Genetic Diversity of Arabusta coffee and their parental genotypes (MSc thesis), Kenyatta University, Kenya. p 171. |
|
|
Hindorf H, Omondi CO, (2011) A review of three major fungal diseases of Coffea arabica L. in the rainforests of Ethiopia and progress in breeding for resistance in Kenya. Journal of Advance Research 2(2):109-120. |
|
|
Jaetzold R, Schimidt H, Hornez H, Shisanya C (2006). Farm Management Handbook of Kenya, Vol. II/C: Natural Conditions and Farm Management Information, Central Kenya. Ministry of Agriculture, Nairobi; Kenya. p.573. |
|
|
Kathurima CW (2013). Characterization of the Diversity of Coffee Genotypes in Kenya by Genetic, Biochemical and Beverage Quality Profile, (PhD thesis) Jomo Kenyatta University of Agriculture and Technology, Kenya. P. 240. |
|
|
Kenya National Bureau of Statistics (2019). Economic survey Report. |
|
|
Kim SM, Reinke RF (2019). A novel resistance gene for bacterial blight in rice, Xa43(t) identified by GWAS, confirmed by QTL mapping using a bi-parental population. PLoS ONE 14(2):e0211775. |
|
|
McDonald J (1926). A preliminary account of a disease of green berries in Kenya colony. Transactions of the British mycological Society 11(1-2):145-154. |
|
|
Mekonnen T, Haileselassie T, Tesfaye K (2017). Identification, Mapping and Pyramiding of Genes/Quantitative Trait Loci (QTLs) for Durable Resistance of Crops to Biotic Stresses. Journal of Plant Pathology and Microbiology 8:412. |
|
|
Minai JM, Nyairo N, Mbataru P (2014). Analysis of socio-economic factors affecting the coffee yields of smallholder farmers in Kirinyaga County, Kenya. Journal of Agricultural and Crop Research 2(12):228-235. |
|
|
Moncada MP, Tovar E, Montoya JC, González A, Spindel J, McCouch S (2016). A genetic linkage map of coffee (Coffea arabica L.) and QTL for yield, plant height, and bean size. Tree Genetics and Genomes 12:5. |
|
|
Mtenga JD (2016). Diversity, combining ability and Coffee Berry Disease (colletotrichum kahawae) resistance among Ethiopian and Tanzanian Arabica coffee genotypes, (PhD thesis), Sokoine University of Agriculture, Tanzania. p 117 |
|
|
Omondi CO, Ayiecho PO, Mwang'ombe AW, Hindorf H (2001). Resistance of Coffea arabica cv. Ruiru 11 tested with different isolates of Colletotrichum kahawae, the causal agent of coffee berry disease. Euphytica 121:19-24. |
|
|
Omondi CO, Gichimu BM, Cheserek J, Gimase J (2016). Leveraging germplasm acquisition for Arabica Coffee improvement in Kenya. Journal of Agriculture and Crop Research 4(1):9-16. |
|
|
Pestana KN, Capucho AS, Caixeta ET, de Almeida DP, Zambolim EM, Cruz CD, Zambolim L, Pereira AA, Oliveira ACB, Sakiyama NS (2015). Inheritance study and linkage mapping of resistance loci to |
|
|
Hemileia vastatrix in Híbrido de Timor UFV 443-03. Tree Genetics and Genomes 11:72. |
|
|
Schneider K (2005). Genetic and Physical Mapping; The Handbook of Plant Genome Mapping; WILEY-VCH Verlag GmbH and Co., KGaA, Weinheim, p 22. |
|
|
Sundaram RM, Vishnupriya MR, Laha GS, Rani NS, Srinivasa RP, Balachandran SM, Reddy GA, Sharma NP, Sonti RV (2009). Introduction of bacterial blight resistance into Triguna, a high yielding, mid-early duration rice variety by molecular marker assisted breeding. Biotechnology Journal 4(3):400-407. |
|
|
Van der Vossen HAM, Cook RTA, Murakaru GNW (1976). Breeding for resistance to Coffee Berry Disease caused by Colletotrichum coffeanum Noack sensu Hindorf in Coffea arabica L. I. Methods of pre-selection for resistance. Euphytica 25:733-756. |
|
|
Van der Vossen, HAM, Walyaro DJA (1980). Breeding for resistance to coffee berry disease in coffea arabica L. II. Inheritance of the resistance. Euphytica 29:777-791. |
|
|
Van der Vossen, HAM, Walyaro DJA (1981). Coffee breeding programme in Kenya. A review of progress made since 1971 and a plan of action for the coming years. Kenya Coffee 46:113-130 |
|
|
Van Der Vossen H, Walyaro D (2009). Additional evidence for oligogenic inheritance of durable host resistance to coffee berry disease (Colletotrichum kahawae) in Arabica coffee (Coffea arabica L.). Euphytica 165(1): 105-111. |
|
|
Van der Vossen H, Bertrand B, Charrier A (2015). Next generation variety development for sustainable production of arabica coffee (Coffea arabica L.): a review. Euphytica 204:243. |
|
|
Vieira A, Diniz I, Loureiro A, Pereira AP, Silva MC, Varzea V, Batista D (2019). Aggressiveness profiling of the coffee pathogen Colletotrichum kahawae. Plant Pathology 68:358-368. |
|
|
Zhao GZ, Jiang CM, Liu JX, Chen YM, Yu TQ, Cheng ZQ (2014). Identification and analysis on the rice blast resistance gene Pi-ta in wild rice from Yunnan, China. China Journal of Rice Science 28(6):675-680. |
|
|
Zhou L, Vega FE, Tan H, Lluch RA., Meinhard LW, Fang W, Mischke S, Irish B, Zhang D (2016). Developing Single Nucleotide Polymorphism (SNP) Markers for the Identification of Coffee Germplasm. Tropical Plant Biology 9:82-95. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


