Full Length Research Paper

ABSTRACT
Xochimilco’s lacustrine zone is a network of channels that, along with the chinampas, conform a unique ecosystem which has served as source of aquatic resources. Duckweeds are small free-floating monocotyledon aquatic plants classified as macrophytes that serve as nutrient pumps and reduce eutrophication effects. Recently, there are number of new studies related to the aquatic plant-microbial interaction focused on the direct functional analysis that investigate plant microbe interactions at full biological hierarchy. The aim of this study was to compare the auxin in vitro production efficiency of the endophyte phytobacteria isolated from Lemna gibba L. plants collected nearby the Xochimilco aquatic agrosystem. There were 17 isolates obtained from the L. gibba plants collected in the dry season and 14 isolates for the rainy season. The environmental conditions and seasonal characteristics determined the number and identity of the isolated endophyte phytobacteria in L. gibba plants according to the several apparent differences in the water quality. This work contributes to the knowledge of the phytobacteria diversity in aquatic plants, particularly in Lemnaceae species; here the majority of the isolates have been characterized as higher indole acetic acid producers, recommended as candidates for their use as biofertilizers.
Key words: Plant growth-promoting bacteria, biofertilizers, Lemna gibba, Xochimilco.
INTRODUCTION
MATERIALS AND METHODS
RESULTS AND DISCUSSION
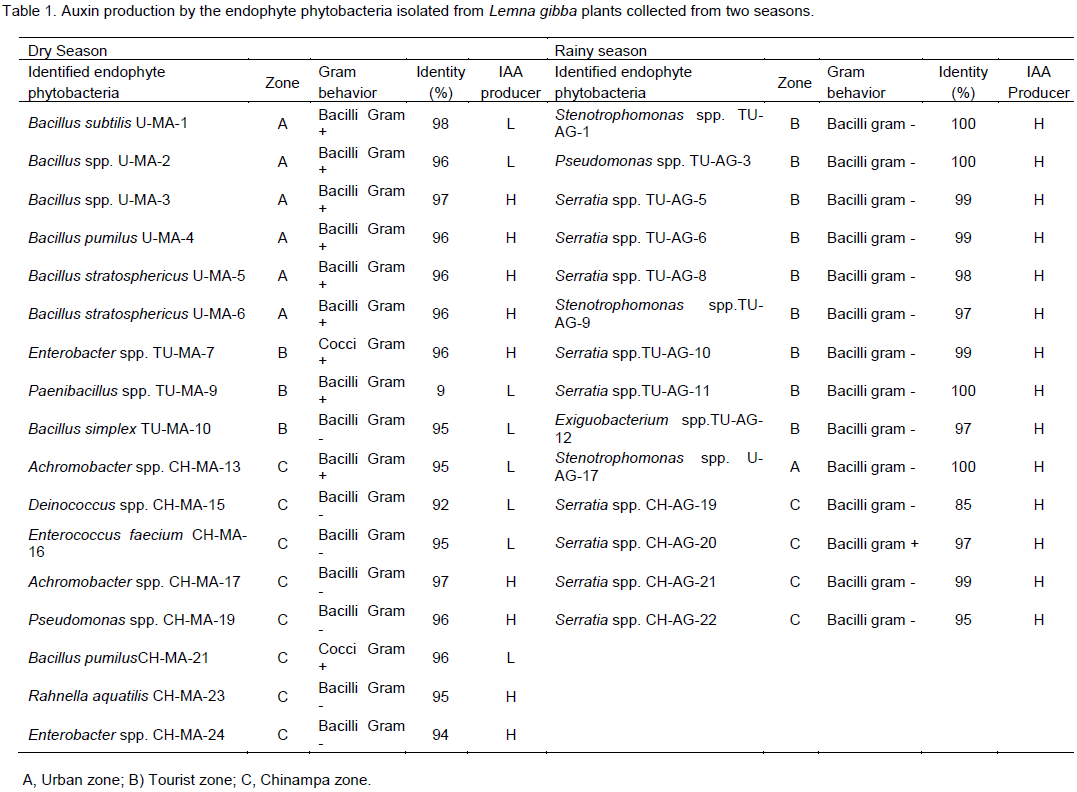
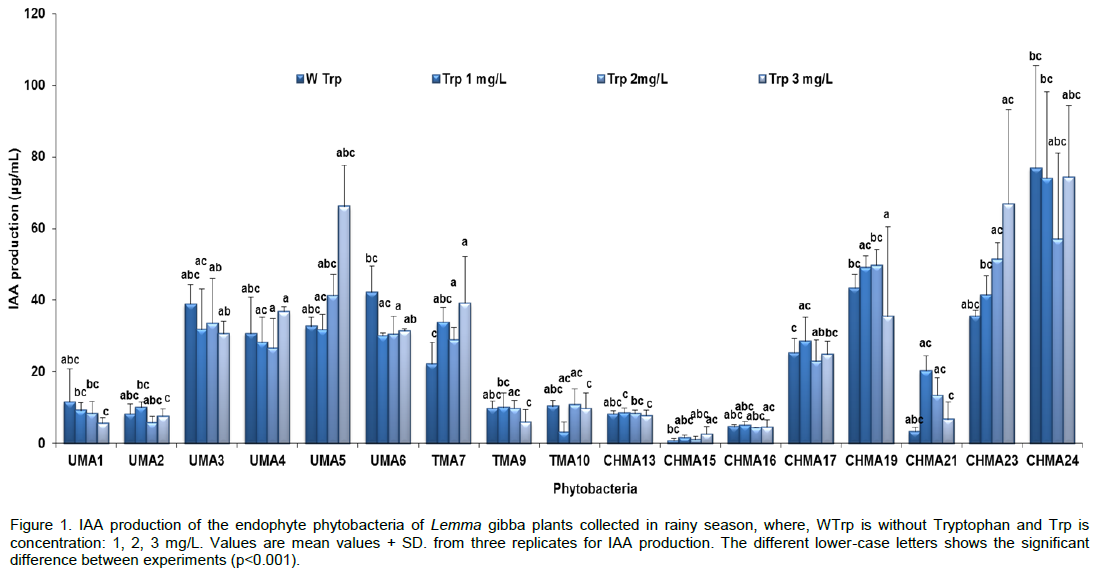
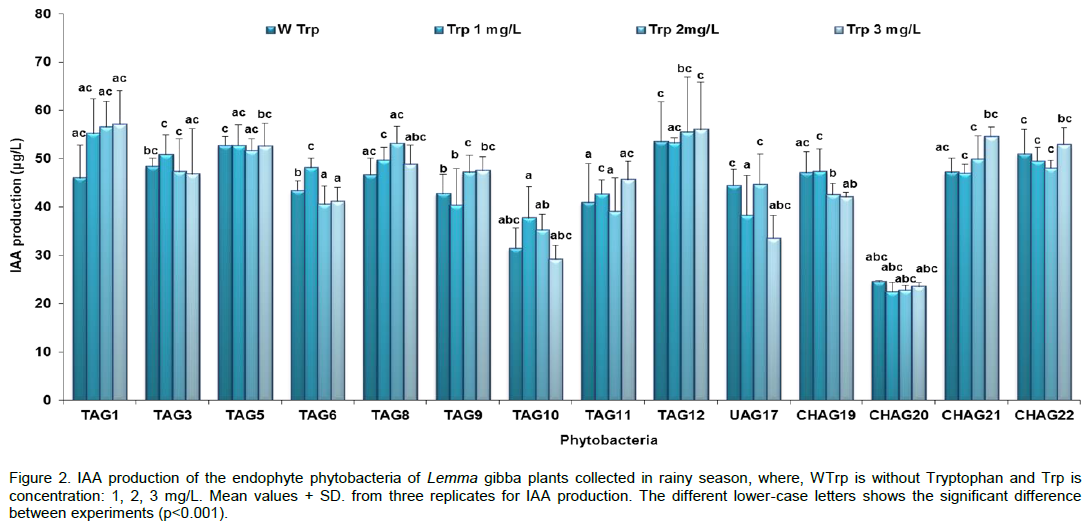
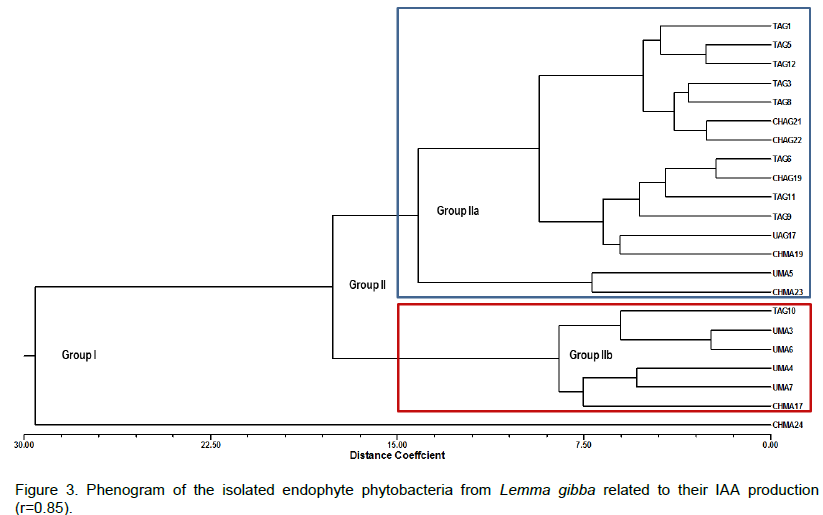
CONCLUSION
This work contributes to the knowledge of the phytobacteria potential in aquatic plants, particularly in Lemnaceae species; here the majority of the isolates have been characterized as higher IAA producers; recommended to candidates for their use as biofertilizers. The authors suggest further works with these isolates that involves plant-microbe systems as biossays to demonstrate the ecological role that these endophyte phytobacteria have in association with L. gibba plants, for the conservation of aquatic ecosystems.
CONFLICT OF INTERESTS
The authors did not declare any conflict of interest.
ACKNOWLEDGMENTS
The authors are grateful to the Research Projects: ISITDF/325/11 AREAS PRIORITARIAS-IPN and SIP-20131494 and SIP-20141314 of the Secretaría de Investigación y Posgrado del I.P.N., for providing the facilities to carry out this work and wish to thank the Comisión de Operaciones y Fomento de Actividades Académicas (COFAA-IPN), EDI (Estímulo al Desempeño de Investigadores-I.P.N.) and Sistema Nacional de Investigadores (SNI-CONACyT), the fellowships for its support throughout the development of this research work.
REFERENCES
|
Allers T, Lichen M (2000). A method for preparing genomic DNA that restrains branch migration of Holiday junctions. Nuclei Acid Res. 28:26-36. |
|
|
Bric JM, Bostock RM, Silversone SE (1991). Rapid in situ assay for indole acetic acid production by bacteria immobilization on a nitrocellulose membrane. Appl. Environ. Microbiol. 57:535-538. |
|
|
Farinati S, DalCorso G, Bona E, Corbella M, Lampis S, Cecconi D, Polati R, Berta G, Vallini G, Furini A (2009). Proteomic analysis of Arabidopsis halleri shoots in response to the heavy metals cadmium and zinc and rhizosphere microorganisms. Proteomics 9:4837-4850. |
|
|
Gagné F, Blaise C, Aoyama LR, Gagnon C, Couillard Y, Campbell P, Salazar M (2002). Biomarker study of a municipal effluent dispersion plume in two species of freshwater mussels. Environ. Toxicol. 17: 149-159. |
|
|
Hasan MR, Chakrabarti R (2009). Use of algae and aquatic macrophytes as feed in small-scale aquaculture: a review. FAO Fisheries and Aquaculture Technical Paper. No. 531. Rome, FAO. 2009. 123p. |
|
|
Hou W, Chen X, Song G, Wang Q, Chi-Chang C (2007). Effects of copper and cadmium on heavy metal polluted waterbody restoration by duckweed (Lemna minor). Plant Physiol. Biochem. 45:62-69. |
|
|
Idris EES, Bochow H, Ross H, Borriss R (2004). Use of Bacillus subtilis as biocontrol agent. VI. Phytohormone like action of culture filtrates prepared from plant growth-promoting Bacillus amyloliquefaciens FZB24, FZB42, FZB45 and Bacillus subtilis FZB37. J. Plant Dis. Prot. 111: 583-597. |
|
|
Idris EES, Iglesias DJ, Talon M, Borriss R (2007). Tryptophan-dependent production of indole-3-acetic Acid (IAA) affects level of plant growth promotion by Bacillus amyloliquefaciens FZB42. Mol. Plant Microbe Interact. 20:619-626. |
|
|
Juárez-Figueroa LA, Silva-Sánchez J, Uribe-Salas FJ, Cifuentes-García E (2003). Microbiological indicators of water quality in the Xochimilco Canals, Mexico City. Salud Pública Méx. 45:389-395. |
|
|
Kamilova F, Kravchenko LV, Shaposhnikov AI, Azarova T, Makarova N, Lugtenberg B (2006). Organic acids, sugars, and L-tryptophane in exudates of vegetables growing on stone wool and their effects on activities of rhizosphere bacteria. Mol. Plant-Microbe Interact. 19:250-256. |
|
|
Khalid A, Arshad M, Zahir ZA (2004).Screening plant-growth-promoting rhizobacteria for improving growth and yield of wheat. J. Appl. Microbiol. 96:473-480. |
|
|
Koga J, Adachi T, Hidaka H (1991). Molecular cloning of the gene for indolepyruvate decarboxylase from Enterobacter cloacae. Mol. Gen. Genet. 226:10-16. |
|
|
Lalke-Porczyk E, Donderski W (2003). Distribution of epiphytic bacteria on the surface of selected species of helophytes and nimpheides from the littoral zone of the southern part of Jeziorak Lake in Poland. Pol. J. Environ. Stud. 12:83-93 |
|
|
Landolt E (1986). The family of Lemnaceae-a monographic study, Vol. 1. Veröffentlichungen des Geobotanischen Institutes der ETH. Stiftung Rübel, Zürich, Switzerland. |
|
|
López-López E, Sede-o-Díaz JE, Favari-Perozzi L (2006). Lipid peroxidation and acetylcholinesterase activity as biomarkers in the black sailfui goodeid, Girardinichthys viviparous (Bustamante) exposed to water from Lake Xochimilco (Mexico). Aquat. Ecosyst. Health 9:379-385. |
|
|
Martin CD, Gerald AM (1994).Nutrient reduction in an in-series constructed wetland system treating landfill leachate. Water Sci. Technol. 29:267-272. |
|
|
Martínez-Cruz P, Hernández-Martínez A, Soto-Castor R, Esquivel-Herrera A, Rangel-Levario J (2006). Use of constructed wetlands for the treatment of water from an experimental channel at Xochimilco, Mexico. Hidrobiológica 16:211-219. |
|
|
Melo MR, Flores NR, Murrieta SV, Tovar AR, Zú-iga AG, Hernández OF, Mendoza AP, Pérez NO, Dorantes AR (2011). Comparative plant growth promoting traits and distribution of rhizobacteria associated with heavy metals in contaminated soils. Int. J. Environ. Sci. Technol. 8:807-816. |
|
|
Niewolak S (1974). Distribution of microorganisms in the waters of the Kortowskie Lake. Pol. Arch. Hydrobiol. 21:315-333. |
|
|
Olah J (1974). Number, biomass and production of planktonic bacteria in the shallow Lake Balaton. Arch. Hydrobiol. 2:193-217. |
|
|
Patten CL, Glick BR (2002). Role of Pseudomonas putida indole-acetic acid in development of the host plant root system. Appl. Environ. Microbiol. 68:3795-3801. |
|
|
Quiroz FA, Miranda MG, Lot A (1982). Uso potencial de algunas hidrófitas como abono verde en la zona chinampera de Xochimilco. Biótica 7:631-633. |
|
|
Rahman MA, Hasegawa H, Ueda K, Maki T, Okumura C, Rahman MM (2007). Arsenic accumulation in duckweed (Spirodela polyrhiza L.): a good option for phytoremediation. Chemosphere 69:493-499. |
|
|
Rajkumar M, Freitas H (2008). Effects of inoculation of plant-growth promoting bacteria on Ni uptake by Indian mustard. Bioresour. Technol. 99:3491-3498. |
|
|
Richardson CJ (1985). Mechanisms controlling phosphorus retention capacity in freshwater wetlands. Science 228:1424-1427. |
|
|
Sharma R, Sharma K, Singh N, Kumar A (2013). Rhizosphere biology of aquatic microbes in order to access their bioremediation potential along with different aquatic macrophytes. Recent Res. Sci. Technol. 5:29-32. |
|
|
Sheng XF, Xia JJ (2006). Improvement of rape (Brassica napus) plant growth and cadmium uptake by cadmium-resistant bacteria. Chemosphere 64:1036-1042. |
|
|
Stout L, Nüsslein K (2010). Biotechnological potential of aquatic plant–microbe interactions. Curr. Opin. Biotechnol. 21:339–345. |
|
|
Stout LM, Dodova EN, Tyson JF, Nüsslein K (2010). Phytoprotective influence of bacteria on growth and cadmium accumulation in the aquatic plant Lemna minor. Water Res. 44: 4970-4979. |
|
|
Stout LM, Nüsslein K (2005). Shifts in rhizoplane communities of aquatic plants after cadmium exposure. Appl. Environ. Microbiol. 71:2484-2492. |
|
|
Suzuki W, Sugawara M, Miwa K, Morikawa M (2014). Plant growth-promoting bacterium Acinetobacter calcoaceticus P23 increases the chlorophyll content of the monocot Lemna minor (duckweed) and the dicot Lactuca sativa (lettuce). J. Biosci. Bioeng. 118:41-44. |
|
|
Uysal Y, Taner F (2009). Effect of pH, temperature, and lead concentration on the bioremoval of lead from water using Lemna minor. Int. J. Phytoremed. 11:591-608. |
|
|
Weisburg WG, Barns SM, Pelletier DA, Lane DJ (1991). 16Ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 173:697-703. |
|
|
Yamaga F, Washio K, Morikawa M (2010). Sustainable biodegradation of phenol by Acinetobacter calcoaceticus P23 isolated from the rhizosphere of duckweed Lemna aoukikusa. Environ. Sci. Technol. 44: 6470-6474. |
|
|
Zaidi S, Usmami S, Singh BR, Musarrat J (2006). Significance of Bacillus subtilis strain SJ-101 as a bioinoculant for concurrent plant grown promotion and nickel accumulation in Brassica juncea. Chemosphere. 64:991-997. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


