Full Length Research Paper

ABSTRACT
An experiment was conducted to determine fatty acid (FA) pro?les in Chinese Maiwa yak milk across the lactation cycle. Sixty-one healthy, similar weight, 4-7 years old, 3-6 parity Maiwa yaks with unrelated background were selected randomly from Hongyuan County, Sichuan province, in southwest of China (with the altitude over 3500 m). The yaks graze on natural pasture all year around without irrigation, fertilizer, or other changes to the pasture. The samples for each animal were collected separately at 1, 2, 3, 4, 5, 6, 7, 15, 30, 60, 120 and 180 days postpartum for fatty acid composition analysis. The most abundant FA species in yak milk were C16:0, C18:1c9, C18:0 and C14:0, all of which varied significantly with the lactation periods. C10:0, C12:0, C14:0 and C16:0 was positively correlated with each other, and negatively correlated with C18:0. C18:2t9c11 was negatively correlated with saturated FA (SFA). C18:1t11, iso-C15:0, C22:0 and C16:1c7 were located in the central positions of the correlation network. C18:1t11 showed close correlation with C18 unsaturated fatty acid (UFA) (C18:1t13 and C18:3n3), C16:0, C19:0, and anteiso-C17:0. Besides with each other, iso-C15:0 and C22:0 were associated with C15:0, anteiso-C15:0, C19:0, C20:0, and C20:0, anteiso-C15:0, iso-C14:0, 9,10-hexyl-C17:0, respectively. C16:1c7 was closely associated with C13:0, C17 FA (C17:0 and 9, 10-hexyl-C17:0), C16:1c9, C20:5n3 and C20:4n6. In conclusion, these data will be informative for the study on the regulatory mechanism of milk FA formation in yaks.
Key words: Correlations, fatty acid profile, lactation periods, Qinghai-Tibetan plateau, yak.
INTRODUCTION
Yak is a species that lives in mountainous areas of Central Asia, mainly found in the highlands of the Nepalese Himalayas, Indian Kashmir, Tibet, Mongolia, and Bhutan (Neupaney et al., 2003; Dhanapati et al., 2003), at an altitude of 2,500 000 to 5,500 000 m (Shi et al., 2019; Zi et al., 2008). The total world population of yaks is estimated at around 14.214.7 million of which China has the largest number of yaks in the world with approximately 13 14 million that comprises > 93.795% of all planet yaks (Sheng-Hua et al., 2013; Dong et al., 2007). Yak milk is a component of the diet in those areas contains 16.5 ± 2.8116.9-17.7% dry matter (DM), 4.90-5.39% protein, 5.53-7.28.8% fat, 4.9 ± 0.61 4.5-5.0% lactose, and 5.4 ± 2.43%0.8-0.9% minerals (Shi et al., 2019; Or-Rashid et al., 2008), thus is considered to be a naturally concentrated milk (Ren et al., 2014), accounting for 15 to 32% of a herder’s daily fat intake (Liu et al., 2011). Considering the importance of fatty acids (FA) in affecting cheese quality, which is the main product of yak milk in Qinghai-Tibetan plateau (Liu et al., 2011; Gottardo et al., 2017), it will be of great value to determinate the FA composition of yak milk.
Milk fat of yak at very high altitudes is richer in polyunsaturated FA (PUFA) (Gottardo et al., 2017; Nikkhah, 2011). The amounts of conjugated linoleic acid (CLA) cis-9, trans-11 in yak cheese has been found to be 4.2 times greater than that in cow cheese (Jiang et al., 2007; Or-Rashid et al., 2008). Eicosapentaenoic acid (C20:5n3, EPA) and docosahexaenoic acid (C22:6n3, DHA)(Gottardo et al., 2017; Liu et al., 2011) are recognized as playing an essential role in human health, and are particularly important for the proper function of the brain, heart, and retina (Simopoulos, 1991; Kris-Etherton et al., 2003; Din et al., 2004; 1991). The greater content of low molecular weight volatile acids (Li et al., 2010) might prevent some human metabolic illness (Williams, 2000; Haenlein, 2004). Although the milk yield of yak has been reported (147 - 487 200-300 kg) and the lactation duration is just 150-200 days daily milk yield is commonly 1.5-2 kg (Dong et al., 2007; Zhou et al., 2014), considering the milk FA composition in each month varies a lot with the change of diet in dairy cow (Palmquist et al., 2006), determining the specific FA profile alteration across the lactation cycle may be beneficial for the utilization of yak milk.
The synthesis of FA are regulated by complex networks (Bionaz and Loor, 2008), results in the close association between different FA compositions (Samková and Kala?, 2021) (Maroteau et al., 2014). Many genes have reported to be responsible for the synthesis of different parts of FA (Stergiadis et al., 2020), such as short and medium chain FA (SMFA) (Zhu et al., 2014), monounsaturated FA (MUFA) (Shi et al., 2013), PUFA (Park et al., 2009) and very long chain FA (VLFA) (Matsuzaka et al., 2007). The milk fatty acids based on genetic and herd parameters was characterized in cow. Although the pathway leading to the formation of FA will not differ between yaks and cows, concentrations in milk fat are probably different. Investigate the exact association among yak milk FA compositions may enhance our understanding about the formation in yaks. The objective of present study was to investigate the FA profile of yak milk across the lactation cycle, and to identify the correlation among yak milk FA compositions. These data may be better for understanding the lactation physiology of yak, and facilitating the utilization of yak milk.
MATERIALS AND METHODS
Animals and sampling
The Animal Care and Use Committee of the Southwest University for Nationalities approved all procedures and experiments (Ministry of Science and Technology, China, revised in June 2004). Sixty-one healthy, similar weight, 4-7 years old, 3-6 parity Maiwa yaks with unrelated background were selected randomly from Hongyuan County, Sichuan province, in southwest of China (with the altitude over 3500 m) (Figure 1). The yaks graze on natural pasture all year around without irrigation, fertilizer, or other changes to the pasture. The experiment was conducted from May to November 2013. The samples for each animal were collected separately at 1, 2, 3, 4, 5, 6, 7, 15, 30, 60, 120 and 180 days postpartum. The yaks were hand milked once daily between 7 am to 8 am. About 100 ml milk samples (divided into 2 tubes after mixture) for each animal were then transferred using ice boxes from the farm to the laboratory in 6 h, and stored at −80°C until analysis.
Fatty acid extraction and analyses
The milk FA were extracted and methyl-esterified protocol was that 2 ml milk was completely mixed with 50 ml chloroform/methanol (2:1) solution (Bligh and Dyer, 1959; Folch et al., 1957). After being centrifuged at 3000 rpm for 5 min, the solution was transferred into a new tube and mixed with 10 ml 20% NaCl following by violent shock. Folch solution (chloroform:methanol:water/3:47:48) was then used for washing the tube wall. After standing for 5 min, the supernatant was removed. A total of 5 ml 0.5 M NaOH-CH3OH was supplemented into the sample for methylation at 60°C for 30 min. Nitrogen gas was then used for evaporating the solution. 5 ml 14% BF3-CH3OH solution was added to the sample following standing at 80°C for 5 min. After the sample was cooled down to room temperature, 8 ml hexane was supplemented and shocked. The saturated NaCl solution was added until hierarchy. Following standing, the supernatant was transferred to a new tube. CuSO4 was used for removing the water, and the supernatant was then used for gas chromatography-mass spectrometry (GC-MS, Agilent 7890A/5970C) analysis equipment with a HP88 column (60 m x 250 μm x 0.2 μm). Approximately 1 μl methylated lipid samples was injected into the GC system with 1:30 split ratio. The temperature of injector was 280°C. The initial temperature of the column was 120°C, which was temperature-programmed at 8°C/min to 145°C, continued to increase to 220°C at 3°C/min and maintained at 220°C for 3 min. The speed of carrier gas was 1 ml/min. The ionization mode was EI, with the ionization energy of 70 eV with 35 to 500 amu mass number. The temperature of interface, ion source and quadrupole rod were 280, 200 and 100°C respectively. The solvent delay was 3 min.
Statistical analyses
The analysis was carried out in duplicates and the data generated was expressed as mean ± standard deviation. Data obtained were statistically analysed by one-way ANOVA using the Statistical Package for the Social Sciences, version 22.0 (SPSS, Chicago, IL, USA), for significant F-statistics. If the overall F-test was significant (P < 0.05), a Fischer’s T-test was performed to significant differences. The correlation analysis among different FA was calculated by Pearson method (2-tailed) using SPSS 22.0. The |r| ≥ 0.65 with P < 0.05 was defined as correlated significantly.
RESULTS
Differences in milk saturated fatty acids across lactation cycle
A total of 38 FA and groups were detected. The FA individuals shorter than C10:0 could not be quantified due to solvent delay. Table 1 shows the fat content and the major FA profile of milk from primiparous yaks across the lactation cycle. The most abundant FA in yak milk were C16:0, C18:1c9, C18:0 and C14:0 across the lactation cycle, consistent with goat, sheep and cow (Markiewicz-K?szycka et al., 2013). In detail, the relative content of C16:0 in yak milk had the highest content level at 2 days, and was decreased in a fluctuation model until 120 days postpartum, however with a sharp increase at 180 days to the similar level as 1 day postpartum. An inverse trend was observed in the relative content of C18:0, which increased in a fluctuation model, peaked at 7 and 120 days postpartum, with a sharp decrease at 180 days. The relative content of C14:0 was continually decreased across the whole lactation periods except a slight increase at 180 days. Considering that the relative content of FA compositions was seriously affected with the decrease of milk yield in the late lactation period, however, different results were observed at 180 days compared with the previous data (except C16:0) (Liu et al., 2011). Although the discrepancy of breeds, grazing model and pasture may result in different FA profile, the exact reason underlying the divergence remains unknown.
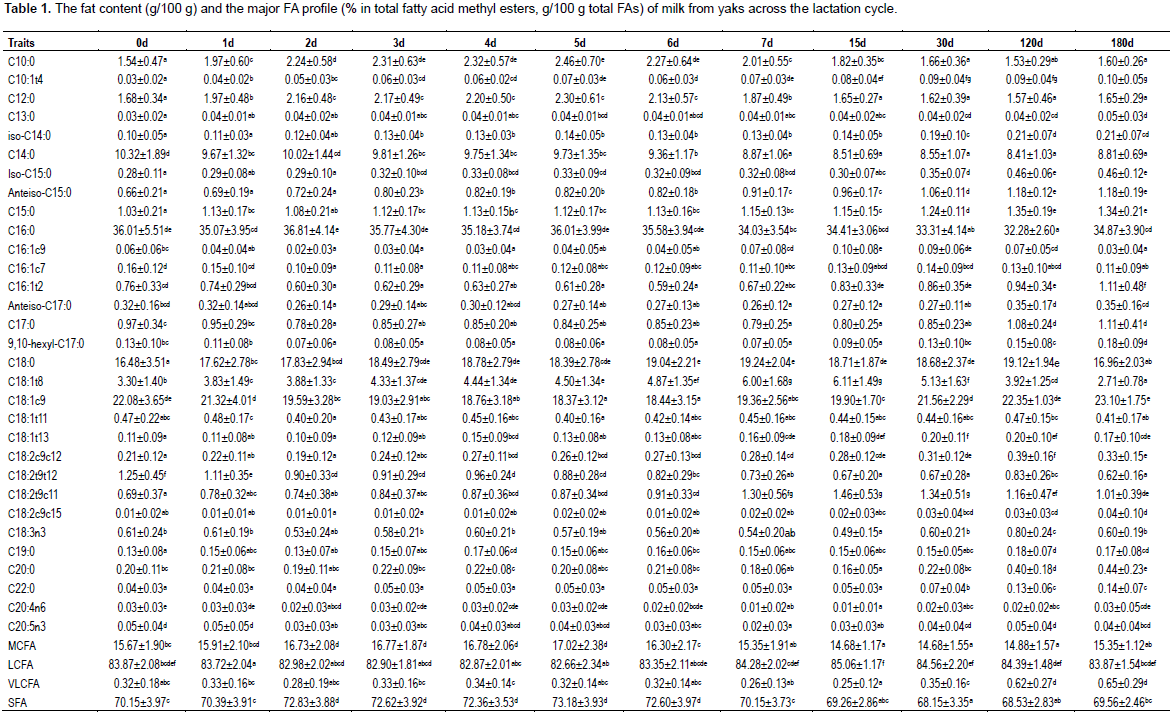

In ruminants, about one-half of milk FA are synthesized de novo (Frutos et al., 2020; Bauman and Davis, 1974), mainly including short- and medium- chain FA (C4-C14) and a portion of 16-carbon FA. Consistent with the data in Gannan yak (Liu et al., 2011), the relative content of C10:0 and C12:0 increased continually until 6 days, and turned down till the end of the lactation cycle. Following a stable period, the relative content of C17:0 was increased significantly at 120 days, however, with no significant change in 9,10-hexyl-C17:0, which was proved to increase cyclooxygenase activity in human. For the LCFA, no significant change was observed in C20:0 and C22:0 except for the significant increase after 30 days postpartum.
In milk fat odd branched-chain fatty acids (OBCFA) are important bioactive component due to their essential role in the gut and potential activity against human breast cancer cells (Gomez-Cortes et al., 2017). In the present study, similar as in dairy cow (Gomez-Cortes et al., 2017), C15:0, C17:0, anteiso-C15:0, iso-C15:0 and anteiso-C17:0 are most abundant OBCFA in yak milk fat. Differently, while a continue increase trend was observed in the content of C15:0 across the lactation cycle, the content of C17:0 decreased continually during colostrum period and increased continually till the end of lactation. However, we did not find iso-C17:0 in the present study. Besides, we also found 9, 10-hexyl-C17:0, the main cyclopropane FA in human adipose tissue and serum, in yak milk, with a continually decrease during colostrum period and turned up till the end of lactation. This may indicate the variation of cyclooxygenase activity during lactation in yaks (Sledzinski et al., 2013).
Differences in milk unsaturated fatty acids across lactation cycle
The C18:2t9t12 was the most abundant FA among the 3 kinds of linoleic acids observed in the present study.
Although there was no significant change for the total content of linoleic acids, C18:2t9t12 continually decreased during lactation periods. Different from our results that cis-9, trans-11 CLA and trans-10, cis-12 CLA were the main CLA in dairy cow (Harvatine et al., 2009; and Wang et al., 2012), dairy goat (Maroteau et al., 2014) and even in Gannan yak (Liu et al., 2011). A possible reason was that the GC system was not sufficient for distinguishing different CLA types only using individual cis-9, trans-11 CLA and trans-10, cis-12 CLA references in yak milk (Liu et al., 2011). In the present study, a GC-MS system was used for more reliable results by comparing the different types of CLA with the NIST mass spectra library (2009). Obviously, more research by GC-MSMS in yak milk is essential for confirming the results in the future.
Consistent with previous study in goat (Haile et al., 2016), dairy cow (Chilliard and Ferlay, 2004) and Gannan yak (Liu et al., 2011), the C18:1c9 was the most abundant USFA. Different with the fluctuation model of C16:0 and C18:0, the relative content of C18:1c9 was continually decreased during colostrum period and turned over after 5 days postpartum. Similar change model was observed in C16:1c9 and C16:1t2 which was the most abundant derivative of palmitic acid. In ruminants, stearoyl-coenzyme A desaturase (SCD) is a primary candidate gene for altering the proportion of SFA versus MUFA in milk with the palmitic acid and stearic acid as substrates (Shi et al., 2013). The relative content of MUFA C18:1c9, compared to the sum of C16:0 and C18:0, was significantly lower than that in dairy goat (Haile et al., 2016) and dairy cow (Cortes et al., 2010), may indicate a lower activity of SCD in yak milk. However, in Gannan yak the value of MUFA/SFA was much higher than that in dairy cow (Wang et al., 2012). Although different breeds of yak may be different in terms of milk fatty acid profile, obviously more controlled study is still needed for elucidating the discrepancy. We also observed small portion of ARA and EPA which presented the activity of fatty acid desaturase 1 (FADS1) (Park et al., 2009), bottomed at 3 and 9 /8 days postpartum, respectively.
Correlations between milk FA
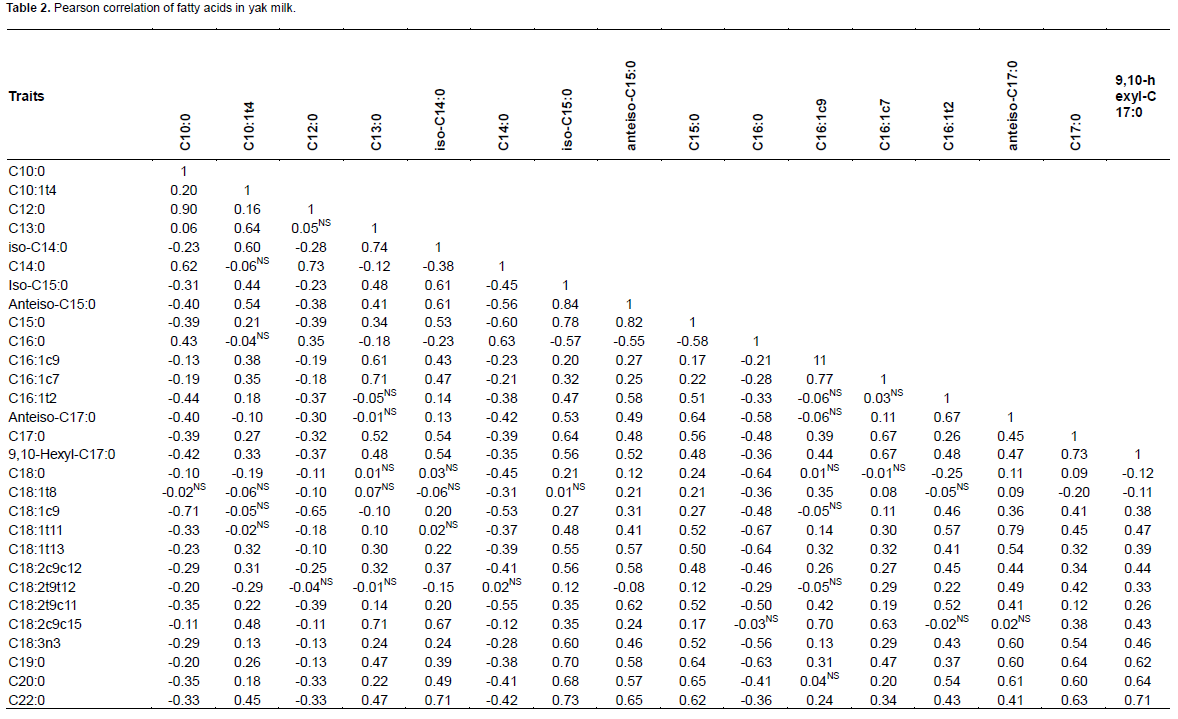
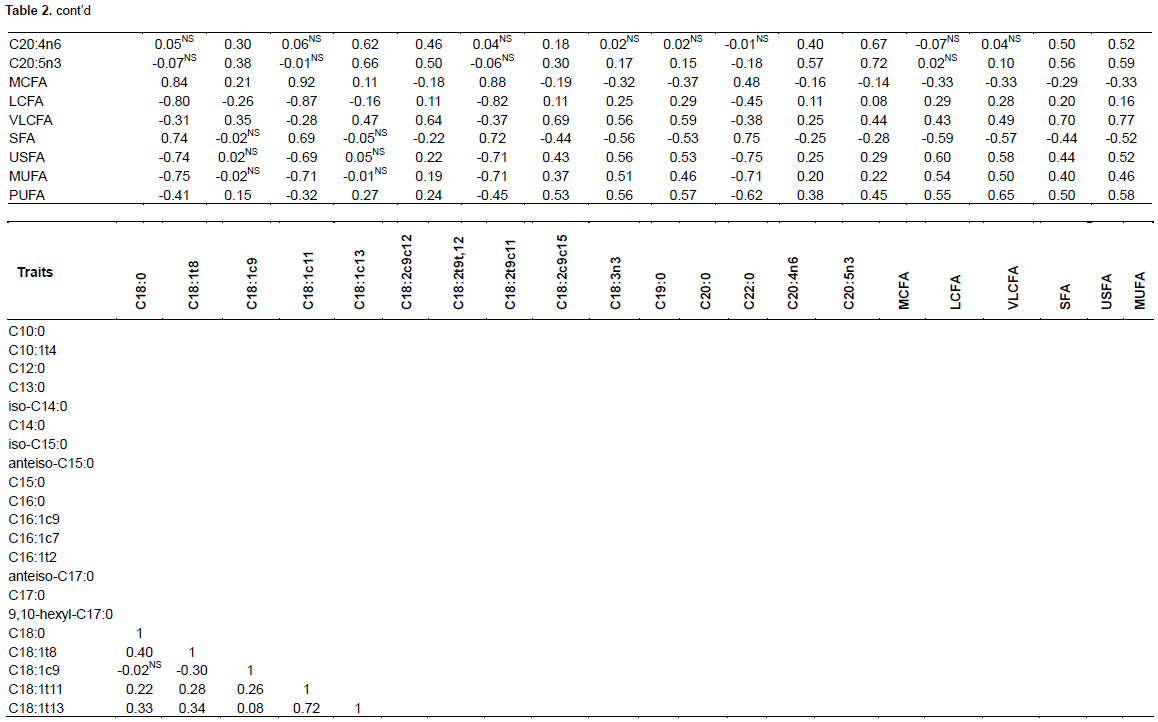
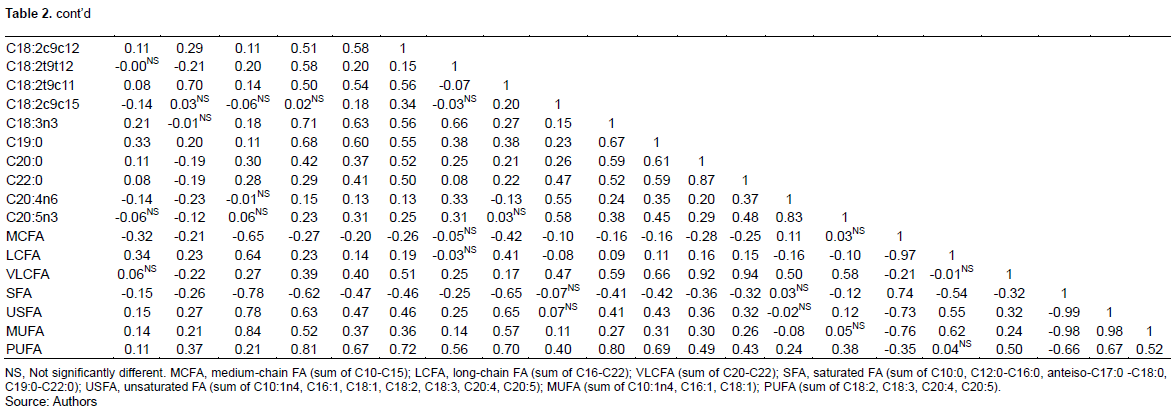
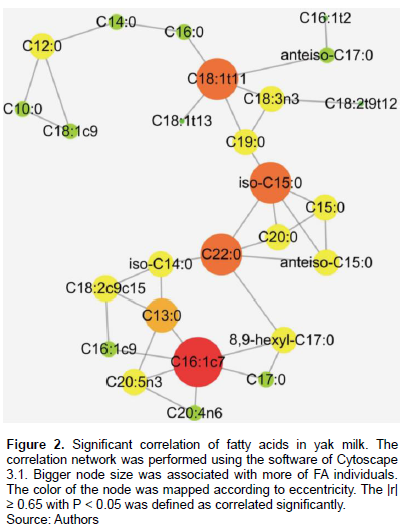
Table 2 showed the FA correlations in Maiwa yak milk, while the Figures 1 and 2 showed the significant correlations. In general, C16:1c7, C18:1t11, C22:0 and iso-C15:0 correlated with most of the FA individuals. As expected, the FA de novo synthesized in mammary gland, including C10:0, C12:0, C14:0 and part of C16:0, significantly correlated with each other, and negatively correlated with C18:0 (r=-0.64). Close correlation was observed among C18 FAs, including C18:1t11, C18:1t13, C18:2t9t12, ALA, and also anteiso-C17:0 (Ntambi and Miyazaki, 2004). While the C16:1c7 was then converted to C18:1c7, C18:1c9 was mainly desaturated from C18:0 (Green et al., 2010). Although C18:2t9c11 was not correlated with the individuals, there was a significantly negative correlation with the total content of SFA, supporting the results of MFD effect on de novo fatty acid synthesis (Piperova et al., 2000).
Close correlations were observed among C15 FAs, C20:0 and C22:0, indicating that anteiso-C15:0, iso-C15:0 and C15:0 may have the same source (Gomez-Cortes et al., 2017). The close correlation between iso-C15:0, as well as 9, 10-hexyl-C17:0 and C17:0, and the total content of VLCFA may indicate their involvement in FA elongation. The C22:0 may be the elongation product of C20:0 in yak milk. Iso-C14:0 was correlated with C13:0 instead of C14:0, indicating that they may share similar source in yak milk (Gomez-Cortes et al., 2017).

DISCUSSION
Although the fatty acid profile of yak milk from the Qinghai-Tibetan plateau in different seasons has been identified (Liu et al., 2011), the present study is novel in that we investigated the variation of milk FA profile across the whole lactation cycle, and analyzed the correlation between FA individuals in grazed yak without the transfer from summer pasture to winter pasture. This was predicted to be the main reason for the discrepancy between our results and Gannan yaks (Liu et al., 2011). In addition, the innovative use of GC-MS, instead of the GC equipment (Or-Rashid et al., 2008; Liu et al., 2011), guaranteed the believable qualitative results of FA composition in yak milk. Although more confirming studies remain needed, these data will be informative for the study of milk FA profile in the future.
We also recognized the limitation of relative quantitative of FA composition. However, this method was used for milk FA profile study in goat (Haile et al., 2016). We also had investigated the variation trend of yak milk FA composition across the whole lactation cycle, and built the correlation among different FA individuals based on a relative quantitative method, which were the main objectives in the present study. Although an absolute quantitative method may be beneficial for detecting the exact content of FA individuals in yak milk, our data also will be informative for understanding the yak milk FA formation regulation.
Another limitation was that we gave a possible that SCD and EVOVLs may be involved in the correlation of FA desaturation and elongation, especially for MUFA and VLCFA, we did not detect the expression of these genes. Obviously, if we could detect the expression of the genes associated with milk fat formation in the mammary gland (Zhu et al., 2014, 2015, 2016), and build their correlation with the FA individuals will enhance our understanding about the regulatory mechanism of FA profile formation in yak milk in the future.
CONCLUSION
This work studied the variation of milk FA profile across the lactation cycle in yak milk grazed at 3,500 m above sea level on the Qinghai-Tibetan plateau. The most abundant FA in yak milk are C16:0, C18:1c9, C18:0 and C14:0, and are varied significantly along with the lactation periods. Close correlations existed among different FA individuals, in which C18:1t11, iso-C15:0, C22:0 and C16:1c7 are at the central positions, indicating that they may paly important role in FA profile formation in yak milk. These data will be informative for the study about the regulatory mechanism underlying the milk FA formation in yak.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
This project was jointly supported by Innovative Postgraduate Research Master Key Project for Southwest Minzu University (CX2019SZ81) the Sichuan Science and Technology Program (2019YFH0035, 21ZDYF2193, 2021ZDYF2706).
REFERENCES
|
Bauman DE, Davis CL (1974). Biosynthesis of milk fat. In Lactation: A Comprehensive Treatise. (BL larson, VR Smith, eds). New York, US: Academic Press pp. 31-75. |
|
|
Bionaz M, Loor J (2008). Gene networks driving bovine milk fat synthesis during the lactation cycle. BMC Genomics 9(1):1-21. |
|
|
Bligh EG, Dyer WJ (1959). A rapid method of total lipid extraction and purification. Canadian Journal of Biochemistry and Physiology 37(8):911-917. |
|
|
Chilliard Y, Ferlay A (2004). Dietary lipids and forages interactions on cow and goat milk fatty acid composition and sensory properties. Reproduction Nutrition Development 44:467-492. |
|
|
Cortes C, da Silva-Kazama DC, Kazama R, Gagnon N, Benchaar C, Santos GT, Zeoula LM, Petit HV (2010). Milk composition, milk fatty acid profile, digestion, and ruminal fermentation in dairy cows fed whole flaxseed and calcium salts of flaxseed oil. Journal of Dairy Science 93:3146-3157. |
|
|
Dhanapati N, Jin-bo K, Makoto I, Kunihiko S (2003). Study on some functional and compositional properties of yak butter lipid. Animal Science Journal 74:391-397. |
|
|
Din JN, Newby DE, Flapan AD (2004). Omega 3 fatty acids and cardiovascular disease--fishing for a natural treatment. British Medical Journal 328(7430):30-35. |
|
|
Dong S, Long R, Kang M (2007). Milking performance of China yak (Bos grunniens): A preliminary report. African journal of agricultural research 2(3):52-57. |
|
|
Folch J, Lees M, Sloane Stanley GH (1957). A simple method for the isolation and purification of total lipids from animal tissues. Journal of Biological Chemistry 226(1):497-509. |
|
|
Frutos P, Hervás G, Natalello A, Luciano G, Fondevila M, Priolo A, Toral PG (2020). Ability of tannins to modulate ruminal lipid metabolism and milk and meat fatty acid profiles. Animal Feed Science and Technology 269:114623. |
|
|
Gomez-Cortes P, Rodriguez-Pino V, Juarez M, de la Fuente MA (2017). Optimization of milk odd and branched-chain fatty acids analysis by gas chromatography using an extremely polar stationary phase. Food Chemistry 231:11-18. |
|
|
Gottardo P, Penasa M, Righi F, Lopez-Villalobos N, Cassandro M, De Marchi M (2017). Fatty acid composition of milk from Holstein-Friesian, Brown Swiss, Simmental and Alpine Grey cows predicted by mid-infrared spectroscopy. Italian Journal of Animal Science 16(3):380-389. |
|
|
Green CD, Ozguden-Akkoc CG, Wang Y, Jump DB, Olson LK (2010). Role of fatty acid elongases in determination of de novo synthesized monounsaturated fatty acid species. Journal of Lipid Research 51(7):1871-1877. |
|
|
Haenlein GFW (2004). Goat milk in human nutrition. Small Ruminant Research 51(2):155-163. |
|
|
Haile AB, Zhang W, Wang W, Yang D, Yi Y, Luo J (2016). Fatty acid synthase (FASN) gene polymorphism and early lactation milk fat composition in Xinong Saanen goats. Small Ruminant Research 138:1-11. |
|
|
Harvatine KJ, Boisclair YR, Bauman DE (2009). Recent advances in the regulation of milk fat synthesis. Animal: an International Journal of Animal Bioscience 3(1):40-54. |
|
|
Jiang J, Chen S, Ren F, Luo Z, Zeng SS (2007). Yak Milk Casein as a Functional Ingredient: Preparation and Identification of Angiotensin-I-Converting Enzyme Inhibitory Peptides. Journal of Dairy Research 74(1):18-25. |
|
|
Kris-Etherton PM, Harris WS, Appel LJ, Nutrition C (2003). Fish consumption, fish oil, omega-3 fatty acids, and cardiovascular disease. Arteriosclerosis, Thrombosis, and Vascular Biology 23:e20-30. |
|
|
Li H, Ma Y, Dong A, Wang J, Li Q, He S, Maubois JL (2010). Protein composition of yak milk. Dairy Science and Technology 90:111-117. |
|
|
Liu HN, Ren FZ, Jiang L, Ma ZL, Qiao HJ, Zeng SS, Gan BZ, Guo HY (2011). Fatty acid profile of yak milk from the Qinghai-Tibetan Plateau in different seasons and for different parities. Journal of Dairy Science 94(4):1724-1731. |
|
|
Markiewicz-K?szycka M, Wójtowski J, Kuczy?ska B, Puppel K, Czy?ak-Runowska G, Bagnicka E, Strza?kowska N, Jó?wik A, Krzy?ewski J (2013). Chemical composition and whey protein fraction of late lactation mares' milk. International Dairy Journal 31(2):62-64. |
|
|
Maroteau C, Palhière I, Larroque H, Clément V, Ferrand M, Tosser-Klopp G, Rupp R (2014). Genetic parameter estimation for major milk fatty acids in Alpine and Saanen primiparous goats. Journal of Dairy Science 97(5):3142-3155. |
|
|
Matsuzaka T, Shimano H, Yahagi N, Kato T, Atsumi A, Yamamoto T, Inoue N, Ishikawa M, Okada S, Ishigaki N, Iwasaki H, Iwasaki Y, Karasawa T, Kumadaki S, Matsui T, Sekiya M, Ohashi K, Hasty AH, Nakagawa Y, Takahashi A, Suzuki H, Yatoh S, Sone H, Toyoshima H, Osuga J, Yamada N (2007). Crucial role of a long-chain fatty acid elongase, Elovl6, in obesity-induced insulin resistance. Nature Medicine 13(10):1193-1202. |
|
|
Neupaney D, KIMJ-b, IshioroshiI M, Samejima K (2003). Study on some functional and compositional properties of yak butter lipid. Animal Science Journal 74(5):391-397. |
|
|
Nikkhah A (2011). Science of Camel and Yak Milks: Human Nutrition and Health Perspectives. Food and Nutrition Sciences 02:667-673. |
|
|
Ntambi JM, Miyazaki M (2004). Regulation of stearoyl-CoA desaturases and role in metabolism. Progress in Lipid Research 43(2):91-104. |
|
|
Or-Rashid MM, Odongo NE, Subedi B, Karki P, McBride BW (2008). Fatty Acid Composition of Yak (Bos grunniens) Cheese Including Conjugated Linoleic Acid and trans-18:1 Fatty Acids. Journal of Agricultural and Food Chemistry 56(5):1654-1660. |
|
|
Palmquist D, Stelwagen K, Robinson P (2006). Modifying milk composition to increase use of dairy products in healthy diets. Animal Feed Science and Technology 131(3-4):149-153. |
|
|
Park WJ, Kothapalli KS, Lawrence P, Tyburczy C, Brenna JT (2009). An alternate pathway to long-chain polyunsaturates: the FADS2 gene product Delta8-desaturates 20:2n-6 and 20:3n-3. Journal of Lipid Research 50(6):1195-1202. |
|
|
Piperova LS, Teter BB, Bruckental I, Sampugna J, Mills SE, Yurawecz MP, Fritsche J, Ku K, Erdman RA (2000). Mammary lipogenic enzyme activity, trans fatty acids and conjugated linoleic acids are altered in lactating dairy cows fed a milk fat-depressing diet. Journal of Nutrition 130(10):2568-2574. |
|
|
Ren QR, Zhang H, Guo HY, Jiang L, Tian M, Ren FZ (2014). Detection of cow milk adulteration in yak milk by ELISA. Journal of Dairy Science 97(10):6000-6006. |
|
|
Sheng-Hua He, Ying Ma, Qi-Ming Li, De-Hai Li, Jiaqi Wang, Yah-Hua Cui (2013). The fatty acid profile of Chinese Maiwa yak milk in relation to season and parity. International Journal of Dairy Technology 66(1):25-30. |
|
|
Samková E, Kala? P (2021). Rapeseed supplements affect propitiously fatty acid composition of cow milk fat: A meta-analysis. Livestock Science 244:104382. |
|
|
Shi B, Jiang Y, Chen Y, Zhao Z, Zhou H, Luo Y, Hu J, Hickford JGH (2019). Variation in the Fatty Acid Synthase Gene (FASN) and Its Association with Milk Traits in Gannan Yaks. Animals 9(9):613. |
|
|
Shi HB, Luo J, Yao DW, Zhu JJ, Xu HF, Shi HP, Loor JJ (2013). Peroxisome proliferator-activated receptor-gamma stimulates the synthesis of monounsaturated fatty acids in dairy goat mammary epithelial cells via the control of stearoyl-coenzyme A desaturase. Journal of Dairy Science 97(12):7844-7853. |
|
|
Simopoulos AP (1991). Omega-3 fatty acids in health and disease and in growth and development. The American Journal of Clinical Nutrition 54(3):438-463. |
|
|
Sledzinski T, Mika A, Stepnowski P, Proczko-Markuszewska M, Kaska L, Stefaniak T, Swierczynski J (2013). Identification of cyclopropaneoctanoic acid 2-hexyl in human adipose tissue and serum. Lipids 48:839-848. |
|
|
Stergiadis S, Cabeza-Luna I, Mora-Ortiz M, Stewart RD, Dewhurst RJ, Humphries DJ, Watson M, Roehe R, Auffret MD (2020). Unravelling the Role of Rumen Microbial Communities, Genes, and Activities on Milk Fatty Acid Profile Using a Combination of Omics Approaches. Frontiers in Microbiology 11:590441. |
|
|
Wang Q, Chang H, Ren Y, Fan E (2012). The difference of fatty acid composition from yak and yellow cattle: Infona Portal Komunikacji Naukowej. |
|
|
Williams C (2000). Dietary fatty acids and human health. Annales de Zootechnie 49:165-180. |
|
|
Zhou ML, Jiang SH, Xie RQ, Luo XL, Yang PG (2014). <Study on the Milk Yield Pattern of Maiwa Yak.pdf>. Animal Husbandry and Feed Science 6(5):261-263. |
|
|
Zhu JJ, Luo J, Sun Y, Shi H, Li J, Wu M, Yu K, Haile AB, Loor JJ (2015). Short communication: Effect of FASN inhibition on triglyceride accumulation and impact on lipid metabolism genes in goat mammary epithelial cells. Journal of Dairy Science 98(5):3485-3491. |
|
|
Zhu JJ, Luo J, Wang W, Yu K, Wang HB, Shi HB, Sun YT, Lin XZ, Li J (2014). Inhibition of FASN reduces the synthesis of medium-chain fatty acids in goat mammary gland. Animal: an International Journal of Animal Bioscience 8(9):1469-1478. |
|
|
Zhu JJ, Luo J, Xu HF, Wang H, Loor JJ (2016). Short communication: Altered expression of specificity protein 1 impairs milk fat synthesis in goat mammary epithelial cells. Journal of Dairy Science 99(6):4893-4898. |
|
|
Zi XD, Lu H, Yin RH, Chen SW (2008). Development of embryos after in vitro fertilization of bovine oocytes with sperm from either yaks (Bos grunniens) or cattle (Bos taurus). Animal Reproduction Science 108(1-2):208-215. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


