Full Length Research Paper

ABSTRACT
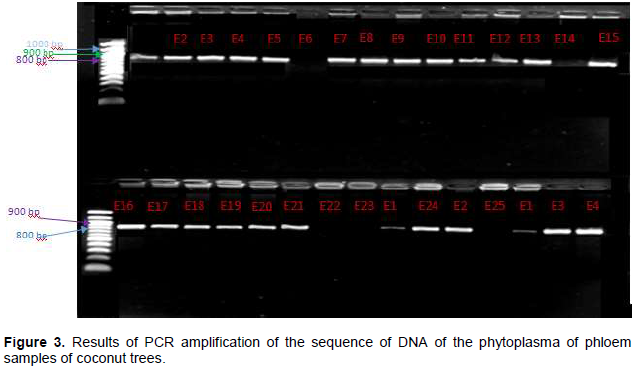
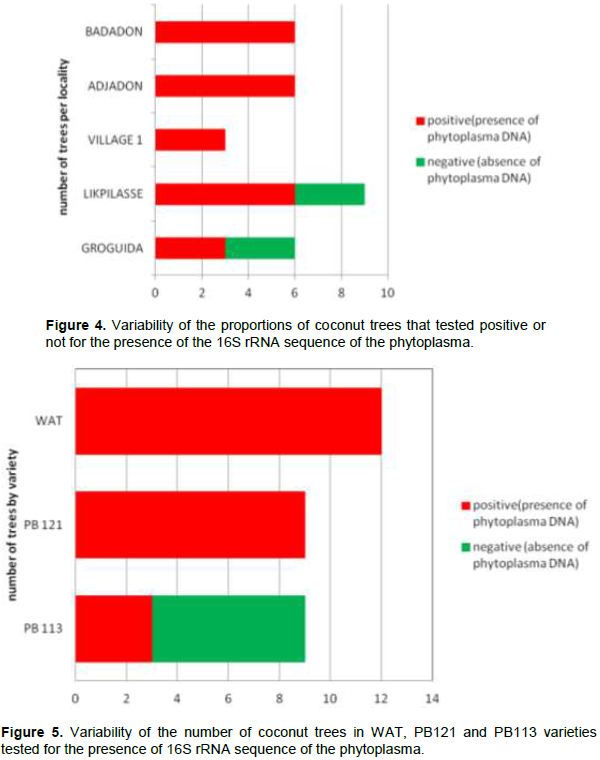
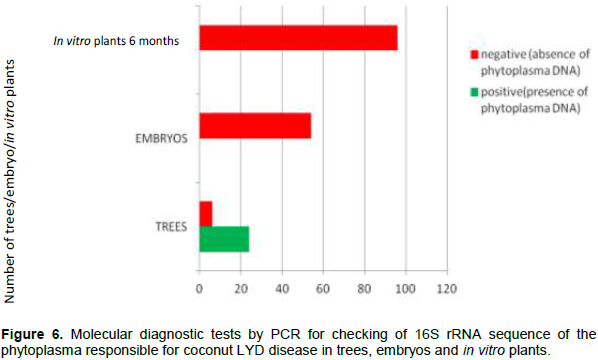
The aim of this study was to investigate the transmission of the lethal yellowing disease (LYD) of coconut tree caused by a phytoplasma from the zygotic embryo to the regenerated plantlet in vitro. From a total of 30 trees, 150 mature coconut nuts where harvested. These nuts were used to extract 150 zygotic embryos. From this package, 96 zygotic embryos were used to regenerate 96 young coconut seedlings in vitro and the 54 others were used to extract total DNA. From the stem of the 30 palms at the stage 1 of the LYD, phloem sample were also collected. From the regenerated in vitro-plantlets at 6 months age, leaf sample were collected. From the molecular diagnosis by PCR, 80% of the phloem samples carried the 16S rRNA gene of the phytoplasma responsible for LYD. All the zygotic embryos and in vitro-plantlets regenerated were healthy. So, coconut zygotic embryos can be used for the safe exchange of genetic material regarding lethal yellowing disease. The regenerated in vitro plantlet are free of disease.
Key words: Coconut, phytoplasma, transmission, in vitro.
INTRODUCTION
MATERIALS AND METHODS


RESULTS
DISCUSSION
CONCLUSION
This study was conducted to check the phytoplasma responsible for the coconut lethal yellowing disease of coconut tree in embryos and seedlings. The results of the molecular diagnosis by PCR revealed that unlike affected trees, the zygotic embryos from the nuts harvested from these trees and seedlings regenerated from the in vitro culture of these zygotic embryos, do not carry the gene responsible for lethal yellowing disease. This is due to the fact that in vitro embryo regeneration generates healthy plants. Thus, the use of embryo for the exchange of plant material even from lethal yellowing disease areas is recommended.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Assy-Bah B, Durand-Gasselin T, Engelmann F, Pannetier C (1989). Culture in vitro d'embryons zygotiques de cocotiers (Cocos nucifera L.): Méthode révisée et simplifiée d'obtention de plants de cocotiers transférables au champ. Oléagineux 44(11):515-523. |
|
|
Baudouin L, Lebrun P (2009). Coconut (Cocos nucifera L.) DNA studies support the hypothesis of ancient Austronesian migration from Southeast Asia to America. Genetic Resources Crop Evolution 56:257-262. |
|
|
Been BO (1981). Observation on field resistance to lethal yellowing in Jamaica. Oléagineu, 36(1):9-11. |
|
|
Cordova I, Jones P, Harrison NA, Oropeza C (2003). In situ PCR detection of phytoplasma DNA in embryos from coconut palms with lethal yellowing disease. Molecular Plant Pathology 4:99-108. |
|
|
Cousin MT (2001). Phytoplasmes et phytoplasmoses: caractéristiques, symptômes et diagnostic. Cahiers d'Agriculture 10:361-376. |
|
|
Cueto CA, Johnson VB, Bourdeix R, Engelmann F, Kembu A, Konan JL, Kouassi Kan M, Rivera RL, Vidhanaarachchi V, Wwise SF (2012). Technical guidelines for the safe movement and duplication of coconut (cocos nucifera l.) germplasm using embryo culture transfer protocols. COGENT; Bioversity International, Montpellier, France. |
|
|
Danso K, Quaicoe R, Oduro V, Dery S, Owusu-Nipah J, Amiteye S, Malaurie B (2009). In vitro germination response and plantlets development of healthy and diseased zygotic embryo of coconut. International Journal of Integrative Biology 7(1):26-31. |
|
|
De Nuce De Lamothe, Wuidart W (1982). L'observation des caracteristiques de developpement vegetatif, de floraison et de production chez le cocotier. Oléagineux 37(6):291-300. |
|
|
De Nuce L, Wuidart W (1979). Les cocotiers grands de Port Bouët (Côte d'Ivoire). 1- Grand Ouest Africain, Grand de Mozambique, Grand de Polynésie, Grand de Malaisie. Oléagineux 34(7):339-349. |
|
|
Deng S, Hiruki C (1991). Amplification of 16S rRNA genes from culturable and non-culturable mollicutes. Journal of Microbiological Methods 14:53-61. |
|
|
Dollet M, Jannot C, Baudouin L, Cirad JO (2009). Le cocotier en Afrique et la maladie du jaunissement mortel OCL 16(2):74-75. |
|
|
Gunn BF, Baudouin L, Olsen KM (2011). Independent origins of cultivated coconut (Cocos nucifera L.) in the old world tropics. PLoS ONE 6, e21143. |
|
|
Harries H, Baudouin L, Cardena R (2004). Floating, boating and introgression: Molecular techniques and the ancestry of coconut palm populations on pacific islands. Ethnobotany Research and Applications 2:37-53. |
|
|
Harrison NA, Richardson PA, Tsaï JH (1995). Detection and diagnosis of Lethal Yellowing: conventional methods and molecular techniques. In: Oropeza C, Howard FW, Ashburner GR (eds.) Lethal Yellowing research and practical aspects, Developments in Plant Pathology, Springer Netherlands, pp. 79-91. |
|
|
Harrison NA, Davis RE, Helmick E (2013). DNA Extraction from Arborescent Monocots and How to Deal with Other Challenging Hosts. In Phytoplasmas: methods and protocols (eds. M. Dickinson and J. Hodgetts), Humana Press, Springer New York Heidelberg Dordrecht London, UK, ISSN 1064-3745, pp. 147-58. |
|
|
Jones P, Tymon AM, Mpunami AA (1999). Detection and diagnosis of African Lethal Yellowing like diseases. In: Oropeza C, Verdeil JL, Ashburner GR, Carde-a R, Santamaría J (eds) Current Advances in Coconut Biotechnology, Kluwer Academic Publishers (Dordrecht/ Boston/London) pp. 197-220. |
|
|
Konan Konan JL, Allou K, Atta Diallo H, Saraka Yao D, Koua B, Kouassi N, Benabid R, Michelutti R, Scott J, Arocha-Rosete Y (2013). First report on the molecular identification of the phytoplasma associated with a lethal Yellowing-type disease of coconut palms in Côte d'Ivoire. New Disease Reports 28:3. |
|
|
Konan Konan JL, Lekadou T, N'goran B, Kouassi N, Gbalou Y. (2013) Project FIRCA (Fonds Interprofessionnel pour la Recherche et le Conseil Agricoles). Cocotier CNRA – COC No. 588. Etude de la maladie du cocotier identifiée dans le Départment de Grand-Lahou, 35p. |
|
|
Mccoy RE, Howard FW, Tsai JH, Donselman HM, Thomas D, Basham HG, Atilano RA, Eskafi FM, Britt L, Collins ME (1983). Lethal Yellowing of palms. University of Florida Agricultural Experiment Stations Bulletin 834, Gainesville, Florida, USA. |
|
|
Myrie W, Oropeza C, Sáenz L, Harrison N, Roca MM (2011). Reliable improved molecular detection of coconut lethal yellowing phytoplasma and reduction of associated disease through field management strategies. Bulletin of Insectology 64:203-204. |
|
|
N'Nan O, Borges M, Konan Konan JL, Hocher V, Verdeil J-L, Tregear J, N'Guetta ASP, Engelmann F, Malaurie B (2012). A simple protocol for cryopreservation of zygotic embryos of ten accessions of coconut (Cocos nucifera L.). In Vitro Cellular and Developmental Biology-Plant 48:160-166. |
|
|
Nipah JO, Jones P, Hodgetts J, Dickinson M (2007). Detection of phytoplasma DNA in embryos from coconut palms in Ghana, and kernels from maize in Peru. Bulletin of Insectology, 60:385-386. |
|
|
N'Nan-Alla Oulo, Ernest Gonedele BI, Abou Bakari Kouassi, Auguste Kouassi, Rene Philippe, Bernard Malaurie, Michel Dollet (2014). Isolation of phytoplasma dna from the coconut palms (Cocos nucifera L.) collected from Ghana, Volume – 2(5) pp. 596-500. |
|
|
Oropeza C, Escamilla JA, Mora G, Zizumbo D, Harrison NA (2005). Coconut lethal yellowing. In Coconut genetic resources, edited by Batugal p, Ramanatha Rao V, et Olivier J, IPGRI, Selangor Darul Ehsan (Malaisie), pp. 349-363. |
|
|
Orozco-Segovia A, Batis Ana I, Rojas-Arechiga M, Mendoza A (2003). Seed biology of palms: a review. Palm 45(2):79-94. |
|
|
Rillo EP, Paloma MBF (1991). Storage and transport of zygotic embryos of Cocos nucifera L. for in vitro culture. Plant Genetic Resource Newsletter 86:1-4 |
|
|
Schneider B, Seemüller E, Smart CD, Kirkpatrick BC. 1995. Phylogenetic classification of plant pathogenetic mycoplasmalike organisms or phytoplasmas. In: Razin S, Tully JG, eds. Molecular and Diagnostic Procedures in Mycoplasmology, Volume I. San Diego, CA, USA: Academic Press, 369-79. |
|
|
Tymon AM, Jones P, Harrison NA. 1998. Phylogenetic relationships of coconut phytoplasmas and the development of specific oligonucleotide PCR primers. Annals of Applied Biology 132:437-452. |
|
|
Weintraub PG, Beanland L (2006). Insect vectors of phytoplasmas. Annual Review of Entomology 51:91-111. |
|
|
Yaima Arocha-Rosete., Konan Konan JL, Diallo AH, Allou K, Scott JA. (2014) Identification and molecular characterization of the phytoplasma associated with a lethal yellowing-type disease of coconut in Côte d'Ivoire. Canadian Journal of Plant Pathology 36:141-150. |
|
|
Yoboue K, Alla-N'Nan O, Konan JL, Sie RS, Kouassi M, Yao SD, Koffi BE (2014). Méthode simple d'échange de germoplasme de cocotier (Cocos nucifera L.) par l'utilisation d'embryons zygotiques. Journal of Applied Biosciences 80:7048-7059. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


