Full Length Research Paper

ABSTRACT
INTRODUCTION
Abiotic stresses such as high salinity, drought, extreme temperature and flooding are the main causes of crop loss worldwide, causing a reduction of more than 50% in the average crop yield, particularly in developing countries (Bray et al., 2000). Because plants are sessile organisms, they are continuously exposed to environmental changes. Plants have evolved effective mechanisms to reduce possible damage (Loredana et al., 2011).
Salinity is considered a universal problem that affects approximately 20% of global irrigated cultivated land (Flowers and Yeo, 1995). A survey conducted by FAO indicated that more than 800 million hectares of land are affected by salt worldwide (FAO, 2008). This amount is equal to more than 6% of the world’s total land area (Munns and Tester, 2008). Extreme salinity is a critical environmental factor that inimically affects large agricultural land areas. Plant growth, physiological processes and metabolic processes are all affected (Magome et al., 2008; Zhang et al., 2009). High salt levels cause ionic stress in the form of cellular Cl- accumulation and especially Na+ ion accumulation. Salt stress also changes the homeostasis of other ions such as Ca2+, K+, and NO3- (Loredana et al., 2011).
Roselle (Hibiscus sabdariffa L.) is an important annual herbaceous shrub belonging to the family Malvaceae that is locally known as “karkade” and grows well in tropical and sub-tropical climates (Cobley and Steele, 1976). Roselle may have been domesticated in western Sudan before 4000 BC (Wilson and Menzel, 1964).
No information is available about possible signal transduction pathways related to abiotic stress in Roselle roots. There are two aspects of salt stress: osmotic stress and ionic stress. As soon as plants as subjected to salt stress, osmotic stress had a major effect on water regulation; continuous ionic stress and osmotic stress reduces plant productivity. To acquire salt tolerance, plants must adapt to both stresses (Ueda et al., 2002).
In this study, differential display reverse transcriptase PCR (DDRT-PCR) was used to identify and isolate salt-induced transcripts from Roselle roots under salt stress. A number of salt stress-responsive transcripts were isolated that had not been previously reported in association with salt (NaCl) stress, providing a prelimi-nary step for identifying and characterizing novel gene(s) with regard to their regulatory elements to provide an understanding of plant adaptations to salt stress conditions.
MATERIALS AND METHODS
Plant material and stress treatment
Roselle (Hibiscus sabdariffa L.) seeds were obtained from the Agricultural Research Corporation's Obeid Research Substation in Sudan. Plants were grown in composite soil (peat, sand, soil, 1:1:1) at the Centre of Excellence in Molecular Biology (CEMB) greenhouse at 30°C ± 2 under relative humidity greater than 40%.
A concentrated NaCl treatment was given to the seedlings to their molecular responses to salt stress could be monitored. After 24 h, root samples from treated and control plants were collected, immediately immersed in liquid nitrogen and stored at -80°C for further use (Mohamed et al., 2013). Total RNA was isolated from frozen root samples using a modified CTAB method (Muoki et al., 2012). Agarose gel electrophoresis was used to test the integrity and purity of the total RNA (Figure 1).

Differential display reverse transcriptase polymerase chain reaction (DDRT-PCR)
Total RNA was isolated, and 2 µg of RNA was reverse-transcribed with an anchored oligo-dT primer by using a RevertAid H- First Strand cDNA Synthesis Kit (Fermentas, USA). cDNAs from different samples were stored at -20°C for differential display PCR amplifications.
A total of 9 anchored and 9 arbitrary primers were used (Table 1). A DDRT-PCR reaction was performed in a 25 µL volume containing 0.5 µL of Taq Polymerase (Invitrogen), 1 µL of arbitrary primer, 1 µL of anchored primer, 0.05 mMdNTPs, 2.5 µL of 10× PCR buffer, 1 µL of cDNA, and 1 µL of MgCl2. The cycling conditions were as follows: an initial denaturation at 95°C for 2 min followed by 35 cycles of denaturation at 95°C for 45 s, annealing at 42°C for 1.30 min, extension at 72°C for 45 s, and a final elongation step at 72°C for 10 min.

The10 µL PCR products were separated vertically on denaturing 16% polyacrylamide gels with 5× TBE buffer at 180 V for 150 min and stained with silver (Bassam et al., 1991). A 50 bp DNA ladder was used to estimate the transcript sizes. The polyacrylamide gel was silver-stained according to the Bio-Rad silver stain handbook. The gel was placed in fixation solution (40% ethanol and 10% glacial acetic acid v/v) for 20 min. The gel was then washed 3 times with dH2O and stained with silver staining solution (0.2% AgNO3) for 20 min. The gel was again washed 3 times with dH2O for 20 s, and developing fluid (3% NaOH and 0.05% formaldehyde) was applied for 3-5 min. The gel was visualized by GrabIT v2.5 Trans-illuminator software on a gel documenting system (Ultra-Violet Products). Reamplification and confirmation of the differentially expressed transcripts were also completed.
Cloning and sequence analysis of cDNA fragments
Selected amplified cDNA fragments were excised and extracted from the gel using the crush and soak method employed by Maqbool et al. (2008). Twenty-five microliters of PCR product was reamplified using the same set of primers that generated the differential display product. For each reamplified band, a quality control without reverse transcriptase was added to monitor for potential RNA contamination by residual genomic DNA, which could be amplified during the subsequent PCR. Reamplified PCR products were separated on an agarose gel (Maqbool et al., 2008). Reamplified eluted DNA fragments were cloned into a pCR 2.1 vector with a TA Cloning Kit (Invitrogen, USA), and the vector was transformed into an E. coli strain. Plasmid DNA was isolated from a minimum of five clones per transformation (Bimboim and Doly, 1979), and their aliquots were treated with RNAase A (Sigma, Germany). The positive clones containing inserts were confirmed via a restriction digestion of plasmid DNA with EcoR1 (Fermentas, USA). The digested product was run on a 1.5 % agarose gel at 80 V for 90 min and analyzed under UV light.
DNA Sequencing and data analysis
Sequencing reactions were performed with an ABI Prism Dye Terminator Kit and an ABI model 3100 automated DNA sequencer. Transcripts were sequenced on both strands with M13 primers. The nucleotide sequences or the deduced amino acid sequences of each clone were compared by using ESTs and protein sequences from various databases through the basic local alignment search tool (BLAST) (Altschul et al., 1990).
Semi-quantitative RT-PCR analysis
RT-PCR reactions were performed with pairs of specific primers for each transcript that were designed on the basis of the H. sabdariffa cDNA sequence (Table 2). β-actin was used as a housekeeping control. The thermal cycling conditions were 3 min of initial denaturation at 95°C followed by 40 cycles of denaturation at 95°C for 30 s, annealing at 60°C for 30 s, extension at 72°C for 45 s and a final extension at 72°C for 10 min.

Quantitative real-time RT-PCR
Real-time PCR reactions were carried out in an ABI 7500 system (Applied Biosystems, USA) with Maxima SYBR Green/ROX qPCR Master Mix (2X) (Fermentas, USA). β-actin was used as a housekeeping control to normalize the data. Fifty nanograms of cDNA were used in each reaction. Real-time PCR was programmed for denaturation at 95°C for 3 min and 95°C for 30 s, followed by annealing at 55°C for 30 s, extension at 72°C for 45 s and additional 40 cycles were repeated untilthe annealing phase. Each reaction was performed in triplicate. The relative gene expression study was conducted using SDS v3.1 software (Applied Biosystems, USA).
RESULTS AND DISCUSSION
Differential display is a molecular biology technique described by Liang and Pardee (1992). It is an efficient method for identifying and isolating differentially expressed genes in specific cells or under altered conditions, and it presents numerous advantages: it is fast and saves time; it produces band patterns in 2 days; it is a simple, well-established and widely accessible technique, making it easily applicable for most investigators; in comparison with prior methods, its sensitivity has increased dramatically, resulting in good detection of low-abundance genes; and both induced and repressed genes can be detected and only a small amount of starting material is needed (Liang and Pardee, 1992).
Since 1992, analyzing gene expression has been the primary goal of most molecular biology studies (Liang and Pardee, 1992). In this study, mRNA differential displays and quantitative RT-PCR were used to study Roselle responses to salt stress. The differential display method reveals all aspects of regulation (up and down), as well as the absence/presence of bands to suggest qualitative differences, and it reveals signals with varying intensity, suggesting quantitative differences (Voelckel and Baldwin, 2003).
Nine arbitrary and 9 anchored primer pair combi-nations were used; 9 transcripts were found, and 4 were repressed by salt stress. The banding pattern of cDNA fragments was amplified by the combination of one primer group, as shown in Table 1. To reduce the false positive rate of the mRNA differential display, each primer pair was used to amplify two different sets of RNA isolated from Roselle plants (Lang et al., 2005).
A GenBank database search revealed that four fragments, with sizes ranging from 200-600 bp (Figure 2), presented significant homologies with known genes (Figure 3). The other cDNA fragments showed no homology to known genes. The expression level of five transcripts was evaluated by real-time RT-PCR. The β-actin gene was used as a reference gene to normalize expression levels. The results indicate that all transcripts were overexpressed at different levels in salt-stressed roots compared to the control, with the exception of B1, which was down-regulated. Many salt stress-activated transcripts have been reported in other plant species (Ouyang et al., 2007; Shahid et al., 2012; Wei et al., 2013). The five transcripts exhibiting different expression levels in response to salt stress, transcripts B2 and B3, were similar and showed two-fold higher expression than the control (Figure 4). All of these transcripts were submitted to the NCBI GenBank EST database, and their accession numbers are given in Table 3.
.png)
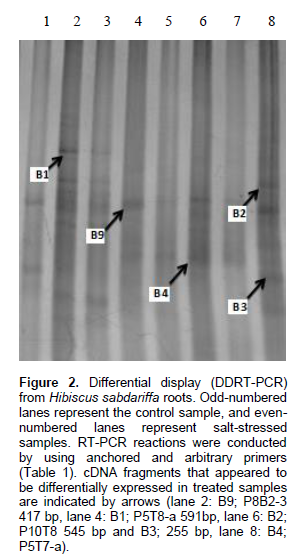
A transcript named Baha-CEMB01 (B1) has high homology with the F-box Arabidopsis protein family (Table 3); it has homology to the Vitis vinifera cultivar Danuta VINE-1 repeat element gag-pol polyprotein. This transcript was down-regulated under real-time PCR.
A transcript named Baha-CEMB02 (B2) has high homology with the F-box Arabidopsis family of proteins (Table 3). The F-box protein family is involved in multiple signaling pathways for regulating root growth; the F-box protein gene reduces abiotic stress tolerance and promotes root growth in rice (Yan et al., 2011). The F-box protein family in eukaryotes plays important roles in plant development and abiotic stress responses via the ubiquitin pathway (Bai et al., 1996; Jia et al., 2011).
Transcripts Baha-CEMB 03 (B3) is homologous with a putative serine/threonine protein kinase (Ipomoea batatas) (Table 3). SAPK4 improved germination, growth and development under salt stress in both seedlings and mature plants. SAPK4-overexpressing rice accumulated less Na+ and Cl- and showed improved photosynthesis in response to salt stress (Diédhiou et al., 2008). In plants, a salt-induced mitogen-activated protein kinase (MAPK) has been identified from alfalfa with a SIMK that is activated by the MAPK kinase SIMKK, and MAPK involvement in osmotic stress signaling has been demonstrated in tobacco and A. thaliana (Sanz, 2003; Slocombe et al., 2002).
The Baha-CEMB 04 (B4) transcript has homology with the putative retrotransposon Ty3-gypsy protein subclass (Oryza sativa Japonica Group). The transcriptional activation of several well-characterized plant retrotrans-posons appears to be tightly linked to molecular pathways activated by stress (Grandbastien, 1998). The Baha-CEMB 09 (B9) transcript has no significant homology with any protein.
Transcripts known as Baha-CEMB02 (B2), Baha-CEMB03 (B3) and Baha-CEMB04 (B4) were identified in Roselle (Hibiscus sabdariffa var. sabdariffa L.), and they shed light on responsive mechanisms to salt acclimation. Identifying these stress-regulated transcripts is an initial step towards cloning and characterizing full-length cDNAs and promoter regions.
This study is the first attempt to investigate the expression levels of roselle transcripts under salt stress by using quantitative real-time PCR. The β-actin gene (DQ866836.1) of Kenaf (Hibiscus cannabinus) actin was used as an internal control. Selected transcripts were overexpressed in the Roselle roots under salt-stressed conditions relative to the control roots (Figures 4 and 5). Transcript B1 was down-regulated, whereas transcripts (B2 and B3) were 1.5-fold higher than those of the control. B4 expression was one-fold higher, and B9 exhibited lower expression. All of these transcripts were submitted to the NCBI GenBank database (Table 3). The regulation of gene expression is one key plant phenomenon used to respond and adapt to salt stress conditions (Jamil et al., 2011).



This study was the first to detect differentially expressed transcripts by applying the differentially expressed mRNA technique to Roselle plants under salt stress and normal conditions. The three identified cDNA transcripts exhibited a higher level of expression under salt stress. These results indicate that Hibiscus sabdariffa variety bulk AlRahad is competent enough to up-regulate some specific genes during salt stress, and these findings are of immense importance for selecting Roselle lines with salt stress-tolerant characteristics.
CONFLICT OF INTERESTS
The author(s) have not declared any conflict of interests.
REFERENCES
|
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990). Basic local alignment search tool. J. Mol. Biol. 215:403-410. Crossref |
||||
|
Bai C, Sen P, Hofmann K, Ma L, Goebl M, Harper JW, Elledge SJ (1996). SKP1 connects cell cycle regulators to the ubiquitin proteolysis machinery through a novel motif, the F-box. Cell 86:263-274. Crossref |
||||
|
Bassam BJ, Caetano-Anollés G, Gresshoff PM (1991). Fast and sensitive silver staining of DNA in polyacrylamide gels. Anal. Biochem. 196:80-83. Crossref |
||||
|
Bimboim HC, Doly J (1979). A rapid alkaline extraction procedure for screening recombinant plasmid DNA. Nucleic Acids Res. 7:1513-1523. Crossref |
||||
| Bray E, BaileyYserres J, Weretilnyk C (2000). Responses to Abiotic Stress, 1158-1203. Biochemistry and Molecular Biology of Plants. American Society of Plant Physiologists, Rockville, USA. | ||||
| Cobley LS, Steele WM (1976). An introduction to the botany of tropical crops, Longman. | ||||
|
Diédhiou C, Popova O, Dietz KJ, Golldack D (2008). The SNF1-type serine-threonine protein kinase SAPK4 regulates stress-responsive gene expression in rice. BMC Plant Biol. 8:49. Crossref |
||||
| Flowers TJ, Yeo AR (1995). Breeding for salinity resistance in crop plants: Where Next? Funct. Plant Biol. 22:875-884. | ||||
|
Grandbastien MA (1998). Activation of plant retrotransposons under stress conditions. Trends Plant Sci. 3:181-187. Crossref |
||||
|
Jamil A, Riaz S, Ashraf M, Foolad MR (2011). Gene Expression Profiling of Plants under Salt Stress. Crit. Rev. Plant Sci. 30:435-458. Crossref |
||||
|
Jia Y, Gu H, Wang X, Chen Q, Shi S, Zhang J, Ma L, Zhang H, Ma H (2011). Molecular cloning and characterization of an F-box family gene CarF-box1 from chickpea (Cicer arietinum L.). Mol. Biol. Rep. 39:2337-2345. Crossref |
||||
|
Lang P, Zhang Ck, Ebel RC, Dane F, Dozier WA (2005). Identification of cold acclimated genes in leaves of Citrus unshiu by mRNA differential display. Gene 359:111-118. Crossref |
||||
|
Liang P, Pardee AB (1992). Differential display of eukaryotic messenger RNA by means of the polymerase chain reaction. Science 257: 967-971. Crossref3 |
||||
| Loredana F, Woodrow P, Fuggi A, Pontecorvo G, Carillo P (2011). Plant Genes for Abiotic Stress. In: "Abiotic Stress in Plants - Mechanisms and Adaptations" (A. Shanker, ed.). InTech. | ||||
|
Magome H, Yamaguchi S, Hanada A, Kamiya Y, Oda K (2008). The DDF1 transcriptional activator upregulates expression of a gibberellin-deactivating gene, GA2ox7, under high-salinity stress in Arabidopsis. Plant J. 56: 613-626. Crossref |
||||
|
Maqbool A, Zahur M, Irfan M, Younas M, Barozai K, Rashid B, Husnain T, Riazuddin S (2008). Identification and expression of six drought responsive transcripts through differential display in desi cottion (Gossypium arboreum). Mol. Biol. 42: 559-565. Crossref |
||||
|
Munns R, Tester M (2008). Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 59:651-681. Crossref |
||||
|
Muoki RC, Paul A, Kumari A, Singh K, Kumar S (2012). An improved protocol for the isolation of RNA from roots of tea (Camellia sinensisL.) O. Kuntze). Mol. Biotechnol. 52: 82-88. Crossref |
||||
|
Ouyang B, Yang T, Li H, Zhang L, Zhang Y, Zhang J, Fei Z, Ye Z (2007). Identification of early salt stress response genes in tomato root by suppression subtractive hybridization and microarray analysis. J. Exp. Bot. 58:507-520. Crossref |
||||
|
Sanz P (2003). Snf1 protein kinase: a key player in the response to cellular stress in yeast. Biochem. Soc. Trans. 31:178-181. Crossref |
||||
| Shahid MN, Jamal A, Rashid B, Aftab B, Husnain T (2012). Identification and isolation of salt-stress-responsive transcripts from Gossypium arboreum L. Turk. J. Biol. 36:746-756. | ||||
|
Slocombe SP, Laurie S, Bertini L, Beaudoin F, Dickinson JR, Halford NG (2002). Identification of SnIP1, a novel protein that interacts with SNF1-related protein kinase (SnRK1). Plant Mol. Biol. 49:31-44. Crossref |
||||
|
Ueda A, Shi W, Nakamura T, Takabe T (2002). Analysis of salt-inducible genes in barley roots by differential display. J. Plant Res. 115:119-130. Crossref |
||||
|
Voelckel C, Baldwin IT (2003). Detecting herbivore-specific transcriptional responses in plants with multiple DDRT-PCR and subtractive library procedures. Physiol. Plant. 118: 240-252. Crossref |
||||
|
Wei S, Wang L, Zhang Y, Huang D (2013). Identification of early response genes to salt stress in roots of melon (Cucumis melo L.) seedlings. Mol. Biol. Rep. 40: 2915-2926. Crossref |
||||
|
Wilson FD, Menzel MY (1964). Kenaf (Hibiscus cannabinus), roselle (Hibiscus sabdariffa). Econ. Bot. 18: 80-91. Crossref |
||||
|
Yan YS, Chen XY, Yang K, Sun ZX, Fu YP, Zhang YM, Fang RX (2011). Overexpression of an F-box protein gene reduces abiotic stress tolerance and promotes root growth in rice. Mol. Plant 4: 190-197. Crossref |
||||
|
Zhang L, Tian LH, Zhao JF, Song Y, Zhang CJ, Guo Y (2009). Identification of an apoplastic protein involved in the initial phase of salt stress response in rice root by two-dimensional electrophoresis. Plant Physiol. 149: 916-928. Crossref |
||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


