Full Length Research Paper

ABSTRACT
Withania somnifera is an important aromatic medicinal plant and possesses wide array of pharmacological properties. In the present investigation, an improved version of hairy root culture system was developed by optimizing various transformation parameters such as type of explant, concentration of acetosyringone, Agrobacterium types and co-cultivation period. Between the leaf and cotyledon explants and two Agrobacterium rhizgenes strains (R1000 and A4) tested, leaf explants infected with R1000 and cocultured for five days on MS basal half strength medium in the presence of acetosyringone (100 µM) attained a higher frequency (88%) of hairy root induction. By adopting this protocol, we could utilize the hairy root culture for industrial scale production of withanolides.
Key words: Leaf explant, Agrobacterium rhizogenes, Withania somnifera, co-cultivation period, acetosyringone.
INTRODUCTION
Withania somnifera (L.) Dunal (Solanaceae), commonly known as ‘ashwagandha’ and “Indian ginseng” is a highly valued medicinal plant in Indian Ayurvedic and African traditional systems. Major withanolides like withanolide A and withaferin A present in W. somnifera have been demonstrated to possess specific therapeutic action against carcinogenesis, Parkinson’s disease and Alzheimer’s disease (Mishra et al., 2000). The requirement of dried plant material for withanolides drugs production in India has been estimated as about 9127-tonnes as against the annual production of about 5905-tonnes (Sharada et al., 2008). Moreover, field cultivation is time consuming, laborious and it is not able to meet the current Ashwagandha global market requirement (Sivanandhan et al., 2012b; 2013a): First, the plant-to-plant variation in secondary metabolites yield along with quality and second the long growing period (4-5 years) between planting and harvesting. To improve commercial cultivation of W. somnifera, biological advances must be made that should either increase the yield or reduce the time gap and assure quality (Banerjee et al., 1994).
To enhance commercial prospects for production of withanolides, an alternative choice could be the use of plant cell/organ cultures. Hairy root cultures offer many advantages over conventional cell culture systems for secondary metabolites production (Georgiev et al., 2007; Sivanandhan et al., 2013b). Hairy roots are adventitious roots derived from cells transformed by the root-inducing plasmid of A. rhizogenes and they grow in the absence of phytohormones. The hairy root harbors the T-DNA segment of plasmid Ri within its genome (Tepfer, 1990). Rao and Ravishankar (2002) suggested A. rhizogenes-mediated hairy root formation as the valuable tool for the biosynthesis of secondary metabolites, metabolic engineering studies and biotechnological production of root-derived compounds.
A. rhizogenes-mediated transformation has been previously reported in W. somnifera using various explants (leaf, cotyledon, cotyledonary node, hypocotyls, stem, and root) and various strains (A4, LBA9402, MTCC 2364, MTCC 532, ATCC 15834, LMG 150, A2/83, A20/83, R1000 and R1601) by various authors (Banerjee et al., 1994; Ray et al., 1996; Pawar and Maheswari, 2004; Kumar et al., 2005; Bandyopadhyay et al., 2007; Murthy et al., 2008; Praveen and Murthy, 2011; Sivanandhan et al., 2012a; Sivanandhan et al., 2013b). In the present study, we documented a higher number of hairy roots and transformation frequency upon inclusion of AS and MES buffer in the co-cultivation medium by optimizing different factors in W. somnifera. Hence, in the present study, two agropine type A. rhizogenes strains (R1000 and A4) have been selected to induce hairy roots in the Kolli Hills genotype of W. somnifera.
MATERIALS AND METHODS
Plant material
The explants, leaf and cotyledon were prepared as per our earlier report (Sivanandhan et al., 2013b).
Agrobacterium rhizogenes-transformation
A single colony of the wild type Agrobacterium rhizogenes strains R1000 and A4 were selected and cultured in LB medium (30 ml; Himedia, Mumbai, India) in darkness at 28°C for 12 h at 180 rpm. The bacterial cells were pelleted by centrifugation followed by washing twice with liquid half strength MS medium (Murashige and Skoog, 1962). The suspension was employed for A. rhizogenes infection.
Standardization of transformation parameters
In order to induce the hairy roots with A. rhizogenes, we tested two types of explants, namely leaves and cotyledons. The explants such as leaf and cotyledon were pricked with sterile hypodermic needle (0.63×25 mm; Dispovan, Haryana, India Ltd.) and the explants were immersed in the bacterial suspension culture (OD 600≥1) for 15 min and blotted on sterile tissue paper for 10 min. After dryness, the explants were placed in half strength MS medium supplemented with different concentrations of acetosyringone (AS) (0, 50, 100, 150, and 200 µM; Sigma, St. Louis, USA), 3% (w/v) sucrose (SRL, Mumbai, India), and 0.2% (w/v) phytagel (Sigma, St. Louis, USA), at 23 ± 2°C in dark conditions. These explants were co-cultivated for different days (3, 5 and 7 days). After the co-cultivation period, the explants were washed first with sterilized-distilled water followed by half strength MS medium, which contained 300 mg/l cefotaxime (Alkime Laboratory, Mumbai, India) and transferred to 30 ml half strength MS medium supplemented with 3% (w/v) sucrose, 0.2% (w/v) phytagel, and 300 mg/l cefotaxime. After 12-18 days of culture, the transformed roots were appeared at the wounded sites of the explants. The induced transformed roots of more than 1-2 cm in length were excised from the explants and transferred to 30 ml half strength MS medium supplemented with 3% (w/v) sucrose, 0.2% (w/v) phytagel and 300 mg/l cefotaxime until the residual bacteria have been completely killed.
Statistical analysis
All the experiments were set up in a completely randomized design and the data were subjected to Duncan's multiple range test using SPSS software version 11.5. The efficiency of hairy root formation was calculated as the percent of respective explants showed hairy root induction out of total number of explants inoculated in a particular treatment.
RESULTS AND DISCUSSION
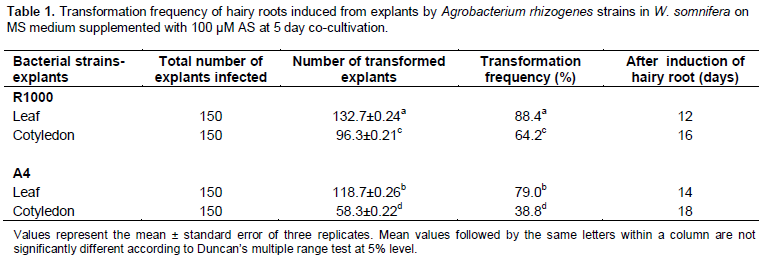
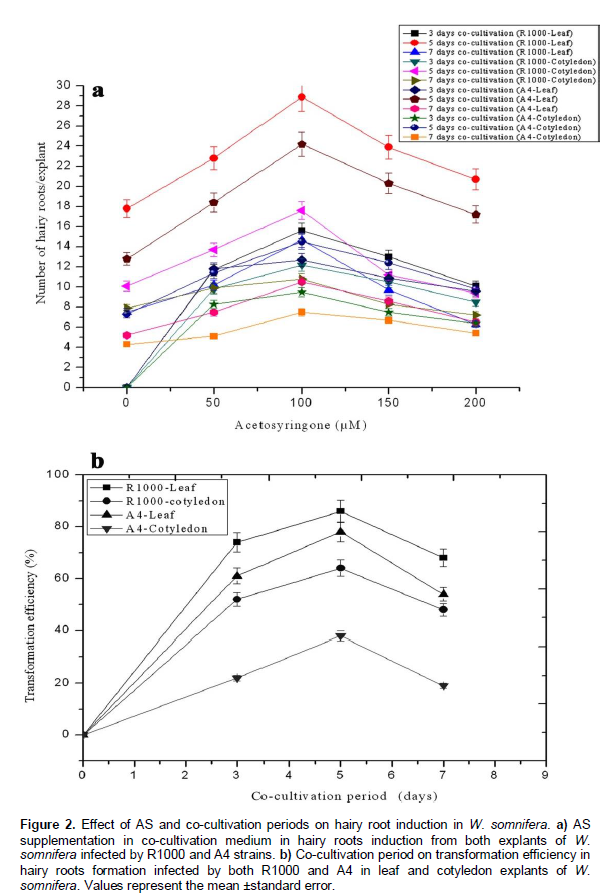
Effect of co-cultivation on hairy root induction
A co-cultivation for 5 days increased the efficiency of the leaf explants with hairy root formation, as it was not observed to obtain a higher number of hairy roots than remaining co-cultivation period. A 5 day co-cultivation period induced 28.2 and 24.3 number of hairy roots/leaf explants infected with R1000 and A4 strains, respectively. In the case of cotyledon explant, the co-culture for 5 days produced 17.6 and 14.5 number of hairy roots/explants infected with R1000 and A4 strains, respectively (Figure 2). Prolonged co-cultivation to 7 days resulted to growth of the bacteria around the both explants which led to explant necrosis. Shorter co-cultivation period may result to an unsuccessful gene transfer from strain to explant. So, the transformation efficiency may differ due to different co-cultivation periods (Figure 2). Of different co-cultivation period tested, 5 day co-cultivation time duration showed dramatic improvement in hairy root induction from the leaf and cotyledon explants, infected with R1000 and A4 strains in W. somnifera. A co-cultivation period for 2 and 3 days was found to be suitable for hairy root induction in W. somnifera, as reported by Pawar and Maheswari, (2004), Murthy et al. (2008), Banerjee et al. (1994), Kumar et al. (2005) and Bandyopadhyay et al. (2007), but Ray et al. (1996) did not mention the co-cultivation duration, whereas in the present study, 5 day co-cultivation period was found to be superior and abundantly increased the efficiency of hairy root induction (Figure 2). These variations in the requirement for a definite co-cultivation period may originate in the specificity of the plant tissue, the Agrobacterium strain or the medium used for bacterial culture and co-cultivation. Co-cultivation is an important phase in transfer of gene from bacteria to plant. But co-cultivation period produced successful transformants on depending upon the plant species and time duration. For R1000 strain, transformation efficiency increased in line with co-cultivation duration and stability on 5th day of culture. During the co-cultivation period, many factors influence the efficiency of the genetic transformation process. After Agrobacterium infection, the plant tissues and bacteria are cultivated for a few days. Important events occur during co-cultivation; plant cells and bacteria divide further, and T-DNA is transferred from the bacteria to the plant cells (Kim et al., 2007). The length of the co-cultivation period also influences transformation efficiency (Tao and Li, 2006). These results confirm that the optimization of co-cultivation period is critical for higher hairy root induction in W. somnifera.
Effect of explants on hairy root induction
Among the various explants tested, only leaf was a greater explant to the infection of both strains. The leaf explant produced higher number of hairy roots by being infected with both the strains R1000 and A4. But the cotyledon explants showed lower number of hairy roots by being infected with both the strains when compared to leaf explants. The leaf explants infected by R1000 and A4 strains produced 28.2 and 24.3 number of hairy roots/explants, respectively and the cotyledon explants infected by R1000 and A4 strains produced 17.6 and 14.5 numbers of hairy roots, respectively (Figure 2). Most of the authors reported that leaf explants only showed the best response for hairy root induction in W. somnifera, but Murthy et al. (2008) obtained lower transformation efficiency in cotyledon explants (3%) whereas in the present study, cotyledon explants also produced hairy roots almost equal to leaf explants. Kang et al. (2006) showed the importance of explant choice by producing more hairy roots of Aralia elata on the root segment than on the petiole explant. It has been reported in several previous studies that the morphological patterns and hairy root production characteristics of infected tissues differed substantially (Ottani et al., 1990). These differences in hairy root production may depend upon differences between species, plant organs, or sites of infection. The choices of explants for hairy root induction after infection with A. rhizogenes constitute the most salient of the integrated factors for a successful hairy root transformation. Plant transformation efficiency differs significantly according to the source of the explant (Alpizar et al., 2006). Therefore, leaf explants possibly influence the rate of hairy root induction than the cotyledon explants by optimizing different factors such as co-cultivation, acetosyringone and bacterial strain in W. somnifera.
ACKNOWLEDGMENTS
Prof. A. Ganapathi is thankful to University Grants Commissions (UGC), Government of India, for the award of UGC-BSR (Basic Scientific Research Fellowship). The first author gratefully acknowledges the Council of Scientific and Industrial Research (CSIR), Government of India, for the award of CSIR-SRF.
CONFLICT OF INTERESTS
The author(s) have not declared any conflict of interest.
REFERENCES
|
Alpizar E, Dechamp E, Espeout S, Royer M, Lecouls AC, Nicole M, Bertrand B, Lashermes P, Etienne H (2006). Efficient production of Agrobacterium rhizogenes-transformed roots and composite plants for studying gene expression in coffee roots. Plant Cell Rep. 25:959-967. Crossref |
||||
|
Bandyopadhyay M, Jha S, Tepfer D (2007). Changes in morphological phenotypes and withanolide composition of Ri-transformed roots of Withania somnifera. Plant Cell Rep. 26:599-609. Crossref |
||||
|
Banerjee S, Naqvi AA, Mandal S, Ahuja PS (1994). Transformation of Withania somnifera (L.) Dunal by Agrobacterium rhizogenes: Infectivity and phytochemical studies. Phytother. Res. 8:452-455. Crossref |
||||
|
Boase MR, Bradley JM, Borst NK (1998). An improved method for transformation of regal pelargonium (Pelargonium Xdomesticum Dubonnet) by Agrobacterium tumefaciens. Plant Sci. 139:59-69. Crossref |
||||
|
Bush AL, Pueppke SG (1991). Cultivar-strain specificity between Chrysanthemum morifolium and Agrobacterium tumefaciens. Physiol. Mol. Plant Pathol. 39:309-323. Crossref |
||||
|
Gelvin SB, Liu CN (1994). Genetic engineering of Agrobacterium tumefaciens strains to improve transformation of recalcitrant plant species. In: Gelvin SB, Schilperoort RA (eds) Plant molecular biology manual. Dordrecht, The Netherlands; Kluwer Academic. pp. 1-13 Crossref |
||||
|
Georgiev MI, Pavlov AI, Bley T (2007). Hairy root type plant in vitro systems as sources of bioactive substances. Appl. Microbiol. Biotechnol. 74:1175-1185. Crossref |
||||
|
Godwin I, Gordon T, Ford-Lloyd B, Newbury HJ (1991). The effects of acetosyringone and pH on Agrobacterium-mediated transformation vary according to plant species. Plant Cell Rep. 9: 671-675. Crossref |
||||
| Kang HJ, Anbazhagan VR, You XL, Moon HK, Yi JS, Choi YE (2006). Production of transgenic Aralia elata regenerated from Agrobacterium rhizogenes-mediated transformed roots. Plant Cell Tissue Org. Cult. 85:187-196. | ||||
|
Kim SJ, Lee JY, Kim YM, Yang SS, Hwang OJ, Hong NJ, Kim KM, Lee HY, Song PS, Kim JI (2007). Agrobacterium-mediated high efficiency transformation of creeping bentgrass with herbicide resistance. J. Plant Biol. 50:577-585. Crossref |
||||
|
Kumar V, Kotamballi N, Chidambara M, Bhamid S, Sudha CG, Ravishankar GA (2005). Genetically modified hairy roots of Withania somnifera Dunal: a potent source of rejuvenating principles. Rejuvenation Res. 8:37-45. Crossref3 |
||||
| Mishra LC, Singh BB, Dagenais S (2000). Scientiï¬c basis for the therapeutic use of Withania somnifera (Ashwagandha): a review. Altern. Med. Rev. 5:334-346. | ||||
|
Murashige T, Skoog F (1962). A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 15:473-497. Crossref |
||||
|
Murthy HN, Dijkstra C, Anthony P, White DA, Davey MR, Power JB, Hahn EJ, Paek KY (2008). Establishment of Withania somnifera hairy root cultures for the production of withanolide A. J. Int. Plant Biol. 50:975-981. Crossref |
||||
|
Nilsson O, Olsson O (1997). Getting to root: the role of the Agrobacterium rhizogenes rol genes in the formation of transformed roots. Physiol. Plant. 100:463-473. Crossref |
||||
| Ottani MP, Schel JHN, Hänisch ten Cate Ch H (1990). Variation in structure and plant regeneration of Agrobacterium rhizogenes transformed and control roots of the potato cv. Bintje. Plant Cell Tissue Org. Cult. 20:25-34. | ||||
| Pawar PK, Maheshwari VL (2004). Agrobacterium rhizogenes mediated hairy root induction in two medicinally important members of family Solanaceae. Indian J. Biotechnol. 3:414-417. | ||||
|
Praveen N, Murthy HN (2011). Synthesis of withanolide A depends on carbon source and medium pH in hairy root cultures of Withania somnifera. Ind. Crop Prod. 35:241-234. Crossref |
||||
|
Rao SR, Ravishankar GA (2002). Plant cell cultures: Chemical factories of secondary metabolites. Biotechnol. Adv. 20:101-153. Crossref |
||||
|
Ray S, Ghosh B, Sen S, Jha S (1996). Withanolide production by root cultures of Withania somnifera transformed with Agrobacterium rhizogenes. Planta Med. 62:571-573. Crossref |
||||
| Sharada M, Ahuja A, Vij SP (2008). Applications of biotechnology in Indian ginseng (ashwagandha): progress and prospects. In: Kumar A, Sopory SK (eds) Recent advances in plant biotechnology and its applications. New Delhi: I. K. International Pvt Ltd. pp. 645-667. | ||||
|
Sivanandhan G, Arun M, Mayavan S, Rajesh M, Jeyaraj M, Kapil Dev G, Manickavasagam M, Selvaraj N, Ganapathi A (2012b). Optimization of elicitation conditions with methyl jasmonate and salicylic acid to improve the productivity of withanolides in the adventitious root culture of Withania somnifera (L.) Dunal. Appl. Biochem. Biotechnol. 168: 681-696. Crossref |
||||
| Sivanandhan G, Kapil Dev G, Jeyaraj M, Rajesh M, Arjunan A, Muthuselvam M, Manickavasagam M, Selvaraj N, Ganapathi A (2013b). Increased production of withanolide A, withanone, and withaferin A in hairy root cultures of Withania somnifera (L.) Dunal elicited with methyl jasmonate and salicylic acid. Plant Cell Tissue Org. Cult. 114:121-129. | ||||
|
Sivanandhan G, Rajesh M, Arun M, Jeyaraj M, Kapil Dev G, Arjunan A, Manickavasagam M, Muthuselvam M, Selvaraj N, Ganapathi A (2013a). Effect of culture conditions, cytokinins, methyl jasmonate and salicylic acid on the biomass accumulation and production of withanolides in multiple shoot culture of Withania somnifera (L.) Dunal using liquid culture. Acta Physiol. Plant. 35: 715-728. Crossref |
||||
| Sivanandhan G, Rajesh M, Arun M, Jeyaraj M, Kapil Dev G, Manickavasagam M, Selvaraj N, Ganapathi A (2012a). Optimization of carbon source for hairy root growth and withaferin A and withanone production in Withania somnifera. Nat. Prod. Commun. 7:1271-1272. | ||||
|
Stachel SE, Messens E, Van Montagu M, Zambryski P (1985). Identification of the signal molecules produced by wounded plant cells that activate T-DNA transfer in Agrobacterium tumefaciens. Nature 318: 624-629. Crossref |
||||
|
Sunikumar G, Vijayachandra K, Veluthambi K (1999). Preincubation of cut tobacco leaf explants promotes Agrobacterium-mediated transformation by increasing vir gene induction. Plant Sci. 141: 51-58. Crossref |
||||
|
Tao J, Li L (2006) .Genetic transformation of Torenia fournieri L. mediated by Agrobacterium rhizogenes. S. Afr. J. Bot. 72:211-216. Crossref |
||||
|
Tepfer D (1990). Genetic transformation using Agrobacterium rhizogenes. Physiol. Plant. 79:140-146 Crossref |
||||
|
Tiwari KK, Trivedi M, Guang ZC, Guo GQ, Zheng GC (2007) .Genetic transformation of Gentiana macrophylla with Agrobacterium rhizogenes : growth and production of secoiridoid glucoside gentiopicroside in transformed hairy root cultures. Plant Cell Rep. 26:199-210. Crossref |
||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


