ABSTRACT
The identification of bee viruses is of great importance where there is no information on their natural occurrence in honeybee populations. Honeybee samples (Apis mellifera L.) obtained from Giza Governorate, Egypt were evaluated for total RNA from the head, thorax and abdomen, and varroa mites. Two fragments of 250 and 540 bp of deformed wing (DWV) and kakugo viruses (KV), respectively, were amplified using conventional real time-polymerase chain reaction (RT-PCR), and were sequenced and analyzed. All bee body parts resulted in a strongly positive DWV sequence except asymptomatic honeybee samples that were negative to the DWV infection. The analysis of varroa mite groups (n = 50) also revealed the presence of DWV which is the first report on this virus in Egypt. Homology search of DWV-PCR amplified fragments revealed 99 and 98% similarities with DWV isolate Warwick-2009 poly-protein gene (accession number: GU109335.1) and the Chilensis DWV isolate, respectively (accession number: JQ413340.1). Also, homology search of KV-PCR amplified fragments using the Genius program 7.1.2 revealed 97 and 90.0% similarities with the KV isolate_Ox genomic sequence (accession number: KC786224.1) and the DWV_Ox genomic sequence (accession number: KC786223.1), respectively. For this study data, a real-time SG RT-Qpcr was found as a fast, simple, sensitive and useful technique to detect and quantify low viral loads in honeybees and varroa mites.
Key words: Deformed wing virus (DWV), kakugo virus (KV), mite, real time-polymerase chain reaction (RT-PCR), Apis mellifera.
The honeybee, Apis mellifera (Hymenoptera: Apidae) is endangered by 18 honeybee single-strand RNA genome monocistronic or dicistronic viruses that have been examined throughout the last decade and cause important economic loses (Allen and Ball, 1996; Anderson, 1991; Bailey and Ball, 1991; Ball, 1997; van Regenmortel et al., 2000; Lanzi et al.,2006). Generally,honeybee viruses are widely distributed and most of them cause asymptomatic and persistent infections (Bailey et al., 1981; Evans and Hung, 2000; Hung and Shimanuki, 1999). Virus infections may be activated by environmental factors (Kukielka et al, 2008b) or parasitic mite infestations which sometimes give rise to the appearance of clinical symptoms. The honeybee viruses exist in association with the mites, Acarapis woodi (Rennie) and Varroa destructor (Anderson and Trueman 2000). Bee colonies infested with large numbers of V. destructor are weakened further by other virus diseases (Smart and Sheppard, 2012).
The deformed wing virus (DWV), the most common among honeybee viruses (Allen and Ball, 1996; Nordstrom et al., 1999), was first isolated in Japan in the 1980s and is now present wherever V. destructor exists (Allen and Ball, 1996; Calderon et al., 2003; Berenyi et al., 2007). DWV infection causes typically deformed wings and other symptoms in drones and workers (De Jong et al., 1982; Schatton-Gadelmayer and Engels, 1988; Bowen-Walker et al., 1999; Nordstrom, 2003). DWV genome sequences have been obtained in most countries (Antunez et al., 2006; Berenyi et al., 2007; Haddad et al., 2008).
The Kakugo virus (KV) is another RNA virus, with a homology sequence very close to two other iflaviruses [DWV and V. destructor virus-1 (VDV-1)]. The virus affects severely, workers’ brains, probably highly correlated with aggressive behavior (Fujiyuki et al., 2004; Lanzi et al., 2006). KV and DWV RNAs have high sequence homology, with 96% nucleotides and 98% amino acid identity, but differ in a few nucleotides (Fujiyuki et al., 2004, 2006).
DWV species complex consists of three distinct variants. DWV type A is classified as two variants, DWV (Lanzi et al., 2006) and Kakugo virus (KV) (Fujiyuki et al., 2004). VDV-1 is now named DWV type B (Martin et al., 2012; Mordacai et al., 2016). DWV-B is found to be more virulent than DWV-A and causes more rapid colony collapse when it infects honey bee colonies (McMahon et al., 2016). DWV-A and B recombinant has been described previously (Moore et al., 2011), and recognized to be hyper virulent than other variants when co-injected to honey bee larvae (Ryabov et al., 2014). Recently, a third novel DWV Type C variant was identified which is clearly related to types A and B (Mordacai et al., 2016).
Electron microscopy, serological methods and others like immunodiffusion, enzyme linked immune-sorbent assay (ELISA), which enhance chemiluminescent Western blotting (Allen et al., 1986; Stolz et al., 1995; Allen and Ball, 1996) are the basis of the classical diagnosis to detect honeybee viruses. DWV and KV are difficult to differentiate due to the high sequence homology between them, but specific molecular techniques have been developed to sensitively detect both genomes (Fujiyuki et al., 2004; Lanzi et al., 2006). The simplicity and sensitivity of the SYBR green-based real-time RT-PCR technique makes it an excellent alternative for detection of honeybee viruses (Kukielka et al., 2008a).
This study aimed to detect honey bee viruses in Egypt and explain the complex relationships between DWV and KV, and also prove that DWV is related to KV by using a simple and sensitive quantitative RT-PCR (RT-qPCR) technique.
Samples collection and RNA extraction
A total number of 300 adult worker bees were collected from 20 collapsing and 10 non-collapsing colonies originating from 10 widely separated apiaries from in Dokki area, Giza governorate, during spring and summer in 2010 to 2012. The bee samples were collected with tweezers randomly from inside the hives. At least, 50 individual varroa mites were also analyzed. Asymptomatic and deformed wing honeybees were collected from the same colonies, and aggressive honeybees (ready to attack according to Fujiyuki et al., 2004) were collected from other colonies and stored immediately at -70°C until total RNA extraction. The specimens were homogenized with 5 mL of PBS solution (pH 7.4). Then, the samples were centrifuged at 1500 rpm for 15 min at 4°C, and the supernatants were collected in sterile Eppendorf tubes, adding BIOZOL-BIOFLUX reagent. The cDNAs were synthesized from the extracted RNAs.
Synthesis of oligonucleotides
Amplification of DWV and KV genomes was performed using two pairs of designed primers to select the polyprotein gene for DWV (Kukielka et al., 2008a) and the RNA dependent RNA polymerase region for KV (Fujiyuki et al., 2004). The nucleotide sequences of DWV primers have accession numbers DQ385507, DQ385505, DQ385502 and NC003784, the Fwd. 5'(CTG TAT GTG GTT GCC TGG T)3' the Rev. 5'(TTC AAA CAA TCC GTG AAT ATA GTG T)3', and the nucleotide sequences of KV primers have accession number AB070959, the Fwd. 5'(ATG TTG CAG CTT CGG CTT TC)3', and Rev. 5'(GGG TCC AAT TCG TTC CT)3'. The specificity of these primers for the viral honeybee infections (Kukielka et al., 2008a) was confirmed using the BLAST® and Clustal W® softwares.
The conventional RT-PCR
The primers were provided by MWG Biotech (Ebensburg, Germany) and Elim Biopharmaceutical, Inc. The cDNA procedure was applied for DWV and KV according to the manufacturer's instructions (Sigma Company- Fermentas, #k1081) in a final 20 µL volume as follows: 7 µL template RNA, 1 µLOligo (dT)18 primer, 4 µL 5X reaction buffer (PH 8.3), 1 µL RibolockTMRNase, 2 µL of 10 mM dNTP mix, 1 µL ReverAidTM H Minus Reverse Transcriptase, and 4 µL DEPC-treated water, and followed by incubation at 43°C for 1 h and was terminated by heating at 70°C for 5 min. The final procedure for the RT-PCR product was performed in a final 50 µL volume using Dream Taq™ Green PCR Master (2X) # K1081, according to Sigma Company Ferments as follows: PCR Dream Taq polymerase master Mix, 2X 25 µL, 1 µL for forward and reverse primers (20 µM), 8 µL template cDNA, and 15 µL nuclease free water. The DWV and KV PCR products were detected with the PCR program as follows: One cycle at 95°C for 3 min for initial denaturation and 35 cycles for 45s at 93°C for denaturation,1min at 61°C reheating for DWV from the thorax and abdomen of deformed wing honeybees, and 55°C for DWV from the head of the deformed wing honeybees and KV from the head of aggressive honeybees, and 1 min at 72°C for extension and finally, 7 min at 72°C for final extension. However, DWV from the head of aggressive honeybees and V. destructor was detected in other conditions as follows: One cycle for initial denaturation at 95°C for 3 min and 35 cycles for 3 min at 93°C for denaturation, 3 min at 40°C for annealing and 2 min at 72°C for extension, and finally 7 min at 72°C for final extension. The RT-PCR products were visualized on1% agarose gel electrophoresis. Two fragments were generated, 250 bp for DWV and 540 bp for KV.
Real-time PCR
Real-time quantitative PCRs were performed in CFX96™ Real-time PCR Detection System using SsoFast™EvaGreen®supermix (Bio-Rad). The PCR program conditions was as follows: an enzyme activation step of 95°C for 10 min followed by 40 cycles of 95°C denaturation for 15 s and reheating at 61°C for 30 s and 72°C for 1 min. Reactions were performed in triplicate in 25 µL using 20 µM of each primer and 2 µL of a 1/20 dilution of cDNA as template. The RT-qPCR products were purified using the Magzina SYBR Green SG qPCR Master Mix. Specificity, sensitivity and reproducibility of the qRT-PCR method were evaluated and compared with the conventional RT-PCR results of the positive samples for KV and DWV. The same samples were proved with the conventional and the SG RT-qPCR, determining their different detection limit according to Kukilka et al. (2008). The CFX Manager™ software (Bio-Rad) was used to analyze the RT-qPCR data.
Data analysis
The sequenced fragment of each virus was analyzed using the BLAST server at the NCBI and Genius 7.1.2 programs. Analytical sensitivity and specificity of the RT-qPCR were evaluated by the efficiency of amplification and melt curve analyses.
Standard curve
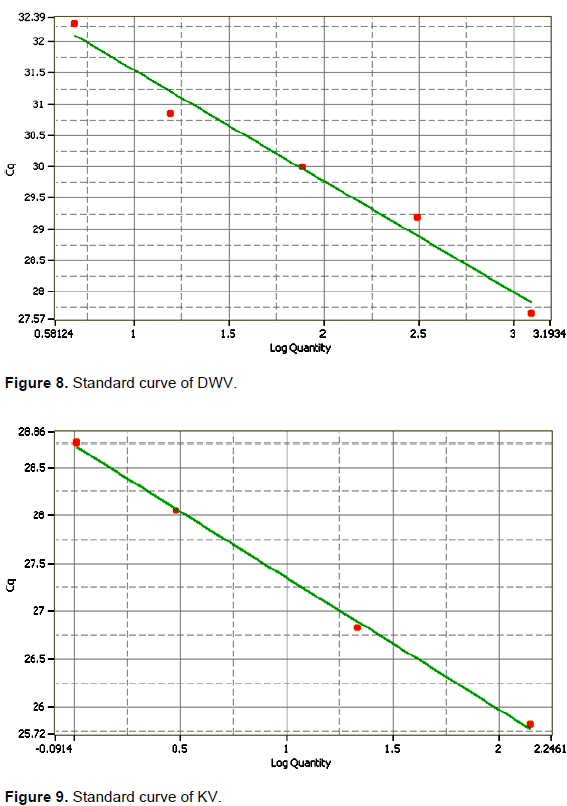
High positive samples for DWV from the thorax and abdomen of deformed honey bees, and KV from the head of aggressive honey bees were used to construct the standard curve, using the sequenced conventional RT-PCR products for each virus as reference (Tencheva et al., 2004). Briefly, serial 4-fold dilutions of these samples were made according to the method by Beuve et al. (2007). The RT-qPCR products were amplified by the same conventional RT-PCR's primers.
Specificity of conventional PCR
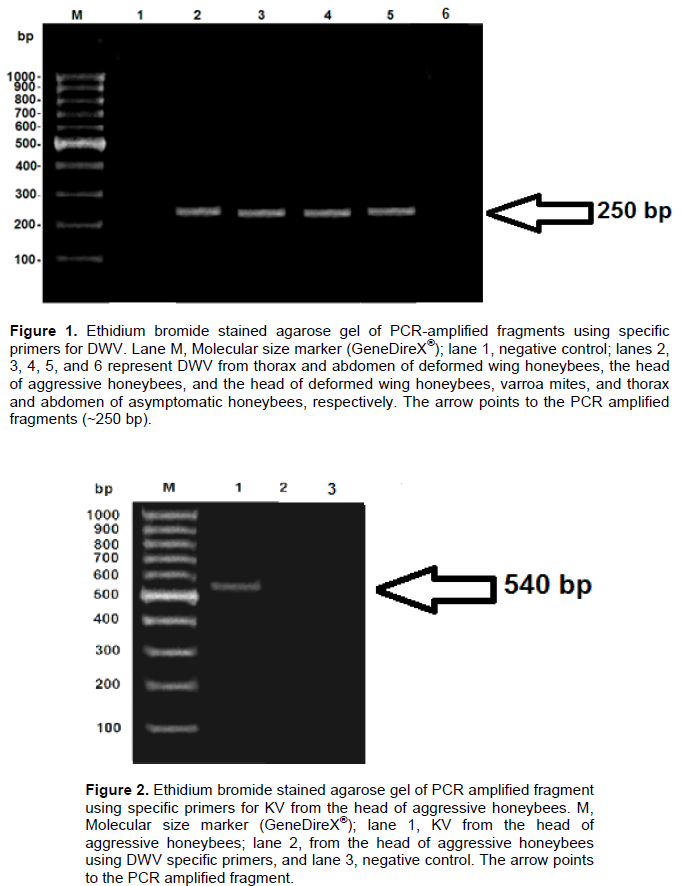
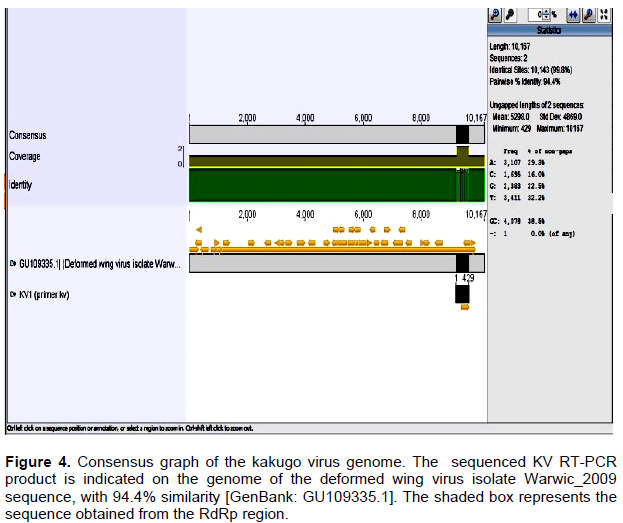
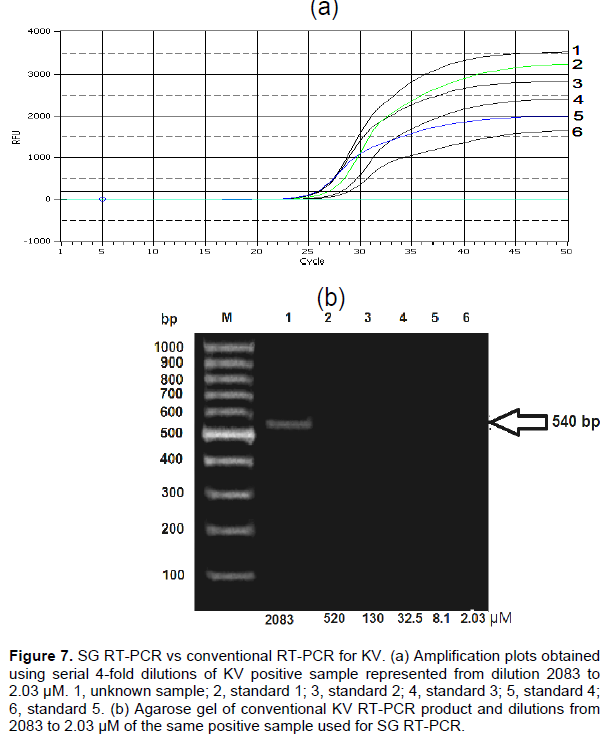
Two fragments (250 and 540 bp) of deformed wing and kakugo viruses, respectively, were amplified using conventional RT-PCR. DWV fragment was generated from head, thorax and abdomen of deformed wing honeybees, head of aggressive honeybees and varroa mites but asymptomatic honeybee samples were negative to DWV infection (Figure 1). KV fragment was generated only from head of aggressive honeybees (Figure 2).
DWV-DNA sequence homologies
Homology search by using the Genius program 7.1.2 revealed DNA similarities of 99% with DWV isolate Warwick-2009 poly-protein gene (accession number GU109335.1) and 98.5% with DWV isolate Chilensis A complete genome (accession number: JQ413340.1) (Figure 3).
KV-DNA sequence homology
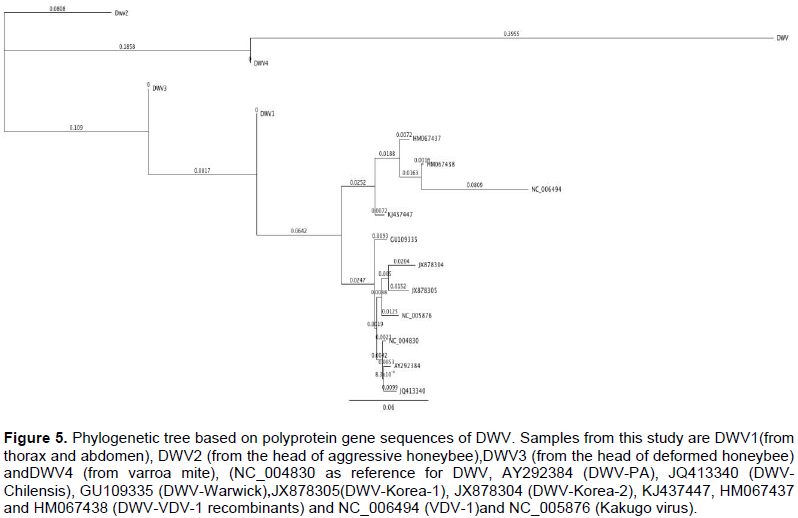
Homology search by using the Genius program 7.1.2 revealed DNA similarities of 97.0% with Kakugo virus isolate KV_Ox genomic sequence (accession number: KC786224.1), 90.9% with DWV_Ox genomic sequence (accession number: KC786223.1) and 94.4% with DWV virus isolate Warwick-2009 poly-protein gene (accession number: GU109335.1) (Figures 3 and 4, respectively).
Phylogenetic analyses of DWV
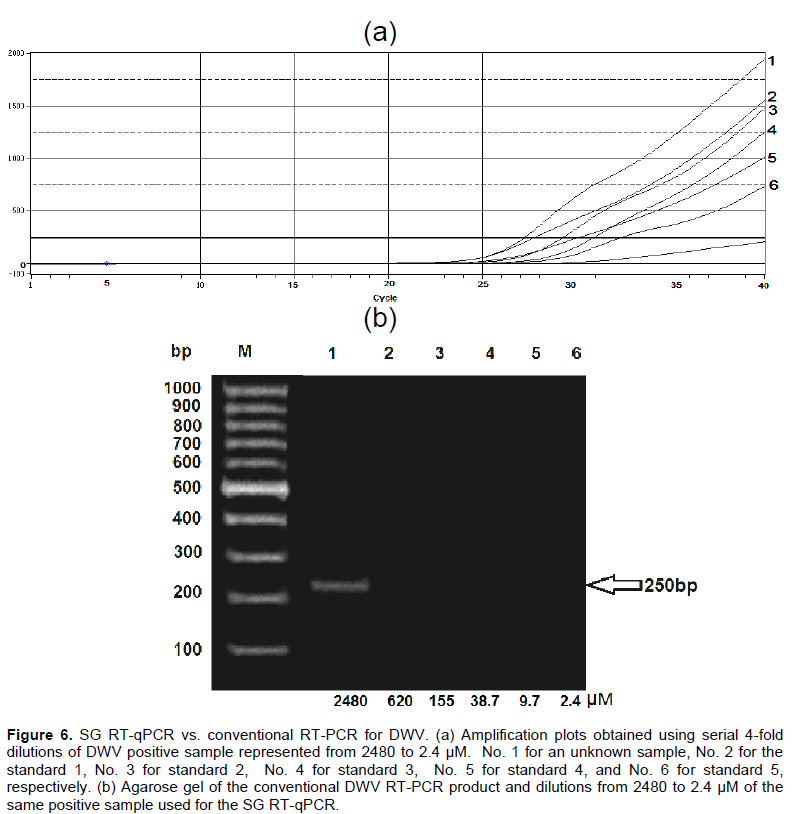
Phylogenetic tree was constructed based on polyprotein gene nucleotide sequences of DWV obtained in this study and 11 related viruses in GenBank to illustrate the genetic relationships among the DWV complex. The amplified fragments were directly sequenced in both directions. They were aggregated and aligned by using the complete DWV sequences deposited in GenBank as references. DWV-VDV-1 recombinants, VDV-1 and Kakugo virus were included in the analysis. The phylogenetic tree indicated that DWV is closely related to KV and clusters together while, DWV-VDV-1 recombinants and VDV-1 appeared to be more close to each other than to DWV and KV and form a distinct clade (Figure 5).
Specificity, sensitivity and reproducibility of the real-time RT-PCR
Alignment of the sequenced PCR amplified fragments of
DWV and KV with the sequences at the GenBank resulted in a sequence identity between 90 and 100% for both viruses. The detection (dilution) limits were 2480 to 2.4 µM, and 2083 to 2.03 µM for DWV and KV (Figures 6 and 7, respectively). For the DWV RT-qPCR, the standard deviation of the mean Cq values obtained from the standard curve ranged from 0.5 to 1.17%, with 0.05 confidence interval ranging from 0.168 to 1.8%. But for DWV from different parts of the honeybee body and the varroa mites, the standard deviation ranged from 0.78 to 1.75%, with 0.05 confidence interval from 22.9 to 3099.6%. For KV, the standard deviation ranged from 0.2 to 0.8% with 0.05 confidence interval from 0.38 to 0.72%.
Standard curve
The slope (S) of the linear regression curve correlated with the efficiency of the PCR reaction was 0.55 and -0.72 where the R2 was 0.977 and 0.99 for DWV from the thorax and abdomen of deformed wing honeybees and KV from the head of aggressive honeybees (Figures 8 and 9, respectively).
Conventional RT-PCR versus real-time RT-PCR
The conventional RT-PCR and RT-qPCR presented different sensitivity.
DWV,which causes a symptomatic chronic infections among bee populations was detected by RT-PCR in the head, thorax and abdomen of infected bees. The transmission of DWV occurs by uncontrolled varroa mites infestations which cause colony collapse and is the main virus associated with varroa (Ball and Allen, 1988; Bailey and Ball, 1991, 1994; Ball and Bailey, 1997; Bowen-Walker et al., 1999; Nordstrom et al., 1999; Nordstrom, 2000; Martin, 2001). Herein, the DWV primer pair yielded a DWV 250 bp fragment that was amplified from the head, thorax and abdomen of deformed wing honeybees, verifying DWV RNA in body sections of infected bees including the head of the aggressive workers. Similar findings were reported by Shah et al. (2009) for A. mellifera in Japan. A 540 bp KV fragment which was amplified from the head of the aggressive workers was generated by using KV specific primers. The specificity of both fragments was confirmed by sequencing and analyzing using NCBI /BLAST server and Genius 7.1.2 programs.
Conventional RT-PCR was performed to detect the presence of DWV and KV in the collected samples. Eight bee viral infections were detected by using several RT- PCR techniques (Soltz et al., 1995; Benjeddou et al., 2001; Grabensteiner et al., 2001; Ribiere et al., 2002; Tencheva et al., 2004; Chen et al., 2005; Genersch, 2005; Yue and Genersch, 2005; Chantawannakul et al., 2006; Maori et al., 2007; Haddad et al., 2008). DWV of honeybees and the recently analyzed KV share 96% nucleotide and 98% amino acid identity (Fujiyuki et al., 2004; Wang et al., 2004). Here in, KV was detected in the head of aggressive bees but not in their thorax and abdomen. Furthermore, the DWV isolated from varroa mites infesting honeybee colonies agrees with the results by Allen et al. (1986) and Bakonyi et al. (2002). RT-PCR and PCR experiments on insects are usually impeded by inhibitor components (Chungue et al., 1993), while SYBR Green (intercalating dye) proved to be more reliable, simple and low cost. Designing the right primers is the principal challenge in performing SYBR-Green qRT-PCR to overcome the formation of the primer dimer, as this dye binds unspecific to any double-stranded (Kukielka et al., 2008a). Herein, two pairs of primers were designed to amplify a fragment of the polyprotein gene of DWV and the non-structural RdRp polyprotein gene of the KV (Berenyi et al., 2007; Terio et al., 2008). Interestingly, these specific primers were defined to be used for qPCR to discriminate the very closely related DWV and KV;Cq 25.6 and Cq 28.05 were obtained when using KV and DWV specific primers, respectively. The amplification plots obtained using serial 4-fold dilutions of DWV 2480 to 2.4 µM from the thorax and abdomen of deformed wing honeybees, the head of aggressive honeybees, the head of deformed wing honeybees and V. destructor revealed that the highest quantity of DWV from the thorax and abdomen of deformed wing honeybees was 2739.1875 µM at Cq 27.16, meanwhile the lowest quantity from the head of deformed wing honeybees was 1.83 µM at Cq 32.91. The quantity of unknown concentration of sample for KV was the highest one (174.55 µM at Cq 25.64). The SG RT-qPCR used here had a detection limit of 9.7 and 1.02 amplicons/µL for DWV and KV, respectively. Therefore, this technique is more sensitive and detects cases of infection more accurately than the conventional RT-PCR. Here, when using the same samples, conventional RT-PCR had a dilution detection limit of 2038 µM for KV and 2480 µM for DWV, while with the SG-based RT-qPCR assay, the dilution detection limit was 2.03 and 9.7 µM for KV and DWV, respectively. Thus, when using the RT-qPCR, viral infections were detected even in samples with a very low viral load, as shown by the detection limit of the RT-qPCR. This high sensitivity and the ability to estimate amplicon copy number in the initial sample allows the detection of unapparent viral infection, avoiding wrong diagnosis.
Within the DWV PCR amplified fragments from all body sections of honeybees and varroa mites, the multiple sequence alignment showed similarity between the four PCR sequences by CLUSTAL W (1.83). Also, it showed 98.5% identity with the JQ413340.1 (Chilensis A deformed wing virus isolate, complete genome) and 99.0% identity with GU109335.1 (Warwick-2009 deformed wing virus isolate) (Figure 3), whereas the sequence alignment of the DWV specific primers used here showed a 3' mismatch with DWV isolate Chilensis A, and had a complete complementary with DWV isolate Warwick-2009 on the GenBank data base by BLASTN and CLUSTAL W. The results are similar to those for the Pennsylvanian and Italian honey bees in which DWV nucleotide sequences were 98% similar to each other and to the (partial) sequences of other DWV isolates (Fujiyuki et al., 2004).
The genetic distance between the nucleotide sequences of the amplified PCR product of deformed wing virus (DWV) detected in honeybees in Egypt, for the Warwick and Chilensis isolates were 0.020 and 0.025, respectively. The 540 bp PCR amplified fragment was 97% identical to KV KC786224.1 (Kakugo virus isolate_Ox genomic sequence) based on the sequenced product, but 90.9% similar to the KC786223.1 (deformed wing virus isolate Ox_genomic sequence), and 94.4% identical to GU109335.1 (deformed wing virus isolate Warwick-2009 poly-protein), confirming that this fragment was the Kakugo virus (KV) because of the high similarity between the DWV and Kakugo virus (Figures 3 and 4). In this study, the analysis of the phylogenetic tree constructed from the sequence of a part of polyprotein gene of DWV concluded that DWV is closely related to KV and clusters together while, DWV-VDV-1 recombinants and VDVl-1 form a distinct clade. DWV-A and -B have been portrayed as having a single quasi-species (Miranda and Genersch, 2010). A mixture of two novel recombinants (VDV-1VVD and VDV-1DVD), and DWV, with levels of the recombinants surpassing that of DWV might have been found in individual bees and mites (Moore et al., 2011). DWV-B with DWV-A recombination and its rise in naive honeybee populations could be of significant ecological and economic importance (McMahon et al., 2016). In addition, DWV Type C variant is distinct from types A and B. Type C had not recently emerged, but rather is an established DWV variant (Mordecai et al., 2016).
Phylogenetically, these viruses might have moved from A. mellifera to wild bee species and genetically as well as geographically, they are in proximity with host species; they likely play an important role in host range of the viruses (Zhang et al., 2012). Phylogenetically, the UPGMA tree of the KV distance showed a complete 0.019 genome difference between DWV and KV, with a 0.013 difference between each other.
Both the DWV and KV for the first time, in honey bees in Egypt were detected using SG RT-qPCR which is a more sensitive technique than conventional RT-PCR. The high similarity between DWV and KV was confirmed. Also, specific primers to be used for qPCR to discriminate DWV and KV were defined. Further molecular studies are needed to understand the antiviral defense mechanisms of honeybees to protect them against infection by varroa mites.
The authors have not declared any conflict of interest.
REFERENCES
|
Allen M, Ball BV (1996). The incidence and world distribution of the honeybee viruses. Bee World 77:141-162.
Crossref
|
|
|
|
Allen MF, Ball BV, White RF, Antoni WJF (1986). The detection of acute paralysis virus in Varroa jacobsoni by the use of a simple in direct ELISA. J. Apic. Res. 25:100-105.
Crossref
|
|
|
|
|
Anderson DL (1991). Kashmir bee virus- a relatively harmless virus of honeybee colonies. Am. bee J. 131:767-770.
|
|
|
|
|
Bailey L, Ball BV (1991). Honeybee pathology. Second ed. Academic Press, London, UK.
|
|
|
|
|
Bailey L, Ball BV (1994). Honeybee viruses. In: R.G. Webster and A. Cranoff (ed.). Encycloped. Of Virol. Academic Press, San Diego, California. Pp. 654-660.
|
|
|
|
|
Bailey L, Ball BV, Perry JN (1981). The prevalence of viruses of honeybee in Britain. Ann. Appl. Biol. 97:109-118.
Crossref
|
|
|
|
|
Bakonyi T, Farkas R, Szendroi A, Dobos-Kovacs M, Rusvai M (2002). Detection of acute bee paralysis virus by RT-PCR in honey bee and Varroa destructor Weld samples: rapid screening of representative Hungarian apiaries. Apidologie 33:63-74.
Crossref
|
|
|
|
|
Ball BV, Allen MF (1988). The prevalence of pathogens in honeybees (Apis mellifera) colonies infested with the parasitic mite Varroa jacobsoni. Ann. Appl. Biol. 113:237-244.
Crossref
|
|
|
|
|
Ball BV (1997). Varroa and viruses. In: Munn, P., and Jones, R. (eds.), Varroa! Fight the mite. International Bee Res. Assoc. Cordiff, UK. Pp. 11-15.
|
|
|
|
|
Ball BV, Bailey L (1997). Viruses. In: Morse R.A., and Flottum, K. (eds.). Honeybee Pest, Predators and Diseases, 3rd ed. Medina, Ohio. pp. 13-31.
|
|
|
|
|
Benjeddou M, Leat N, Allsop PM, Davison S (2001). Detection of acute bee paralysis virus and black queen cell virus from honeybee by reverse transcriptase-PCR. Appl. and Environ. Microbiol. 67(5):169-193.
Crossref
|
|
|
|
|
Berenyi O, Bakonyi T, Derakhshifar I, Koglberger H, Topolska G, Ritter W, Pechhasker H, Nowotny N (2007). Phylogenetic analysis of deformed wing virus genotypes from diverse geographic origins indicates recent global distribution of the virus. Appl. Environ. Microbiol. 73(11):3605-3611.
Crossref
|
|
|
|
|
Beuve M, Semp L, Lemaire O (2007). A sensitive one-step real time RT-PCR method for detecting grapevine leaf roll-associated virus 2 variants in grapevine. J. Virol. Methods 141(2):117-124.
Crossref
|
|
|
|
|
Bowen-Walker PL, Martin SJ, Gunn H (1999). The transmission of deformed wing virus between honeybees (Apis melliferaL.) by the ectoparasitic mite Varroa jacobsoni Oud. J. Invertebr. Pathol. 73:101-106.
Crossref
|
|
|
|
|
Calderon R, Vanveen J, Arce H, Esquivel M (2003). Presence of deformed wing virus and Kashmir bee virus in Africanized honeybee colonies in CostaRica infested with Varroa destructor. Bee World 84:112-116.
Crossref
|
|
|
|
|
Chantawannakul P, Ward L, Boonham N, Brown M (2006). A scientific note on the detection of honeybee viruses using real-time PCR (Taq-Man) in varroa mites collected a Thai honeybee (Apis mellifera) apiary. J. Invert. Pathol. 9:69-73.
Crossref
|
|
|
|
|
Chen YP, Higgins JA, and Feldlaufer MF (2005). Quantitative real-time reverse transcription-PCR analysis of deformed wing virus infection in the honeybee (Apis mellifera L.). Appl. Environ. Microbiol. 71:436-441.
Crossref
|
|
|
|
|
Chungue E, Roche C, Lepnre MF, Barbazan P, Chanteaus S(1993). Ultra rapid, simple, sensitive and economical silica method for extraction of dengue viral RNA from clinical specimens and mosquitoes by reverse-transcriptase chain reaction. J. Med. Virol. 40:142-145.
Crossref
|
|
|
|
|
De Jong D, De Jong PH, Goncalves LS (1982). Weight loss and other damage to developing worker honeybees from infestation with Varroa jacobsoni. J. Apic. Res. 21(3):155-167.
Crossref
|
|
|
|
|
De Miranda JR, Genersch E (2010). Deformed wing virus. J. Invertebr. Pathol. 103:S48-S61.
Crossref
|
|
|
|
|
Evans JD, Hung AC (2000). Molecular phylogenetics and the classification of honeybee viruses. Arch. Virol. 45:2015-2026.
Crossref
|
|
|
|
|
Fujiyuki T, Takeuchi H, Ono M, Ohka S, Sasaki T, Nomoto A, and Kubo T (2004). Novel insect picorna-like virus identified in the brains of aggressive worker honeybees. J. Virol. 78:1093-1100.
Crossref
|
|
|
|
|
Fujiyuki T, Ohka S, Takeuchi H, Ono M, Nomoto A, and Kubo T (2006). Prevalence and phylogeny of kakugo virus, a novel insect picorna-like virus that infects the honeybee (Apis mellifera L.) under various colony conditions. J. Virol. 80:11528-11538.
Crossref
|
|
|
|
|
Genersch E (2005). Development of a rapid and sensitive RT-PCR method for the detection of deformed wing virus, a pathogen of the honeybees (Apis mellifera). Vet. J. 169:121-123.
Crossref
|
|
|
|
|
Grabensteiner E, Ritter MJ, Carter S, Davison H, Pechhacker J, Kolodziejek O, Boeching I, Derakhshifar R, Moosbeckfer E, Licek E, Nowotny N (2001). Sacbrood virus of the honeybee (Apis mellifera): rapid identification and phylogenetic analysis using reverse transcription-PCR. Clin. Diagn. Lab. Immunol. 8:93-104.
Crossref
|
|
|
|
|
Haddad N, Brake M, Migdadi H, de Mirand JR (2008).First detection of honey bee viruses in Jordan by RT-PCR. Jordan J. Agric. Sci. 4(3):242-246.
|
|
|
|
|
Hung ACF, Shimanuki H (1999). A scientific note on the detection of Kashmir bee virus in individual honeybee and Varroa jacobsoni mites. Apidologie 30:353-354.
Crossref
|
|
|
|
|
Kukielka D, Esperón F, Higes M and Sánchez-Vizcaíno JM (2008a). A sensitive one-step real-time RT-PCR method for detection of deformed wing virus and black queen cell virus in honeybee Apis mellifera. J. Virol. Methods 147:275-281.
Crossref
|
|
|
|
|
Kukielka D, Perez AM, Higes M, Bulboa MC, Sánchez-Vizcaíno JM (2008b). Analytical sensitivity and specificity of a RT-PCR for the diagnosis and characterization of the spatial distribution of three Apis mellifera viral diseases in Spain. Apidologie 39:607-617.
Crossref
|
|
|
|
|
Lanzi G, De Miranda JR, Boniotti MB, Cameron CE, Lavazza A, Cappucci L, Camazine SM, Rossi C (2006). Molecular and biological characterization of deformed wing virus of honeybees (Apis mellifera L.). J. Virol. 80:4998-5009.
Crossref
|
|
|
|
|
Maori E, Lavi S, Moze S, Koch R, Gantman Y, Peret ZY, Edelbaum O, Tanne E, Sela I (2007). Isolation and Characterization of Israeli acute paralysis virus, a dicistro virus affecting honeybees in Israel: evidence for diversity due to intra- and inter species recombination. J. Gen. Virol. 88:3428-3438.
Crossref
|
|
|
|
|
Martin SJ (2001). Varroa destructor reproduction during the winter in Apis mellifera colonies in the UK. Exp. Appl. Acarol. 25:321-325.
Crossref
|
|
|
|
|
Martin SJ, Highfield AC, Brettell L, Villalobos EM, Budge GE (2012). Global Honey Bee Viral Landscape Altered by a Parasitic Mite. Science 336:1304–1306.
Crossref
|
|
|
|
|
McMahon DP, Natsopoulou ME, Doublet V, Fürst M, Weging S, Brown MJ, Gogol-Döring A, Paxton RJ (2016). Elevated virulence of an emerging viral genotype as a driver of honeybee loss. Proc. R. Soc. B 283(1833):20160811.
|
|
|
|
|
Mordecai GJ, Wilfert L, Martin S J, JonesI Mand Schroeder DC (2016). Diversity in a honey bee pathogen: first report of a third master variant of the Deformed Wing Virus quasispecies. ISME J.10:1264-1273.
Crossref
|
|
|
|
|
Nordstrom S (2000). Virus infections. PhD thesis. Swedish University of Agriculture Sciences, Uppsala, Sweden.
|
|
|
|
|
Nordstrom S (2003). Distribution of deformed wing virus within honeybees (Apis mellifera) brood cells infested with the ectoparasitic mite Varroa jacobsoni Oud. J. Invertebr. Pathol. 73:101-106.
|
|
|
|
|
Nordstrom S, Fries I, Aarhus A, Hansen H, Karpa S (1999). Virus infections in nandic honeybee colonies with no, low or severe Varroa jacobsoni infestations. Apidologie 30:475-484.
Crossref
|
|
|
|
|
Ribiere M, Triboulot C, Mathieu L, Aurieres C, Faucon JP, Pepin M (2002). Molecular diagnostic of chronic bee paralysis virus infection. Apidologie 33:339-351.
Crossref
|
|
|
|
|
Ryabov EV, Wood GR, Fannon JM, Moore JD, Bull D, Chandler JC, Mead A, Burroughs N, Evans DJ (2014). A Virulent Strain of Deformed Wing Virus (DWV) of Honeybees (Apis mellifera) Prevails after Varroa destructor-Mediated, or In Vitro, Transmission. PLOS Pathog. 10(6):e1004230.
Crossref
|
|
|
|
|
Schatton-Gadelmayer K, Engles W (1988). Blood proteins and body weights of newly-emerged worker honeybees with different levels of parasitization of body mites. Entomol. Gen. 14:93-101.
|
|
|
|
|
Shah KS, Evans EC, Pizzorno MC (2009). Localization of deformed wing virus (DWV) in the brains of the honeybee Apis mellifera Linnaeus. Virol. J. 6(1):182.
Crossref
|
|
|
|
|
Smart MD, Sheppard WS (2012). Nosema ceranae in age cohorts of the western honeybees (Apis mellifera). J. Invertebr. Pathol. 109:148-151.
Crossref
|
|
|
|
|
Stolz D, Shen XR, Boggis C, Sisson G (1995). Molecular diagnosis of Kashmir bee virus infection. J. Apic. Res. 34:153-160.
Crossref
|
|
|
|
|
Tencheva O, Gauther L, Zappulla N, Dainat B, Cousserana F, Colin ME, Bergoin M (2004). Prevalence and seasonal variations of six bee viruses in Apis mellifera L. and Varroa destructor mite populations in France. Appl. and Environ. Microbiol. 70:7185-7191.
Crossref
|
|
|
|
|
Terio V, Martella V, Camero M, Decaro N, Testinic G, Bonerba E, Tantillo G, Buonovogliac C (2008). Detection of a honeybee iflavirus with intermediate characteristics between kakugo virus and deformed wing virus. New Microbiol. 4:439-444.
|
|
|
|
|
van-Regenmortel MHV, Francki RIB, Fauquement CM, Bishop DHL, Wildy P, Fenner F, Mathews REF, Knudson DL, Brown F, Murphy FA, Ghabrial SA, Jarvis AW, Martell GP, Mayo MA, Summers MD, Fauquet CM, Bishop DHL, Carstens EB, Estes MK, Lemon SM, Maniloff J, Mayo MA, McGeoch DJ, Pringle CR, Wickner RB (2000). Virus taxonomy, Seventh report of the international committee on Taxonomy of viruses. San Deigo: Academic Press.
|
|
|
|
|
Wang X, Zhang J, Lu J, Yi F, Liu C, Hu Y (2004). Sequence analysis and genomic organization of a new insect picorna-like virus. Ectropis oblique picorna-like virus, isolated from Ectopis oblique. J. Gen. Virol. 85:1145-1151.
Crossref
|
|
|
|
|
Yue C, Genersch E (2005). RT-PCR analysis of deformed wing virus in honeybees (Apis mellifera) and mites Varroa destructor. J. Gen. Virol. 86:3419-3424.
Crossref
|
|
|
|
|
Zhang X, He SY, Evans JD, Pettis JS, Yin GF, Chen YP (2012). New evidence that deformed wing virus and black queen cell virus are multi-host pathogens. J. Invert. Pathol. 109(1):156-159.
Crossref
|
|
|
|
|
|
|
|