Full Length Research Paper

ABSTRACT
In this study, the effect of three carbon sources (mannitol, minced potato and sucrose), two immobilization substrates (alginate and wood cylinders), and three surfactants (Tween 20, Tween 40 and Tween 80), were evaluated on eritadenine production using shiitake (Lentinula edodes) mycelium under submerged cultivation, in shake flasks within 20 days. Eritadenine and biomass were measured by HPLC and gravimetrically, respectively. Alginate immobilization of mycelium promoted significant enhancement of eritadenine yields of 88 mg/L, compared to the control (8.7 mg/L) and wood immobilization (14.8 mg/L). Likewise, eritadenine yields (72.4 mg/L) were enhanced by adding surfactant tween 20 to the broths in 0.5%, than control (8.7 mg/L) without surfactant. Tween 40 and 80 did not improve eritadenine yields, but both produced better biomass values (superior to 5 g/L) than the control (3.9 g/L). All carbon sources (sucrose, mannitol, mince potato, and glucose as control) produced similar low eritadenine yields, with best results (10.2 mg/L) by sucrose, although glucose produced the best biomass yields of 3.9 g/L. Also, carbon sources and the best biomass values did not show significant effect on eritadenine production. pH values in the best eritadenine yielding fermentations went down from 6 to 3-4, but pH had a low correlation with eritadenine yields. Finally, all data obtained in the present study are useful for optimizing culture conditions, towards industrialization of this important health improver metabolite (eritadenine).
Key words: Eritadenine, shiitake mushroom, submerged culture, immobilization, surfactants.
INTRODUCTION
MATERIALS AND METHODS
RESULTS AND DISCUSSION
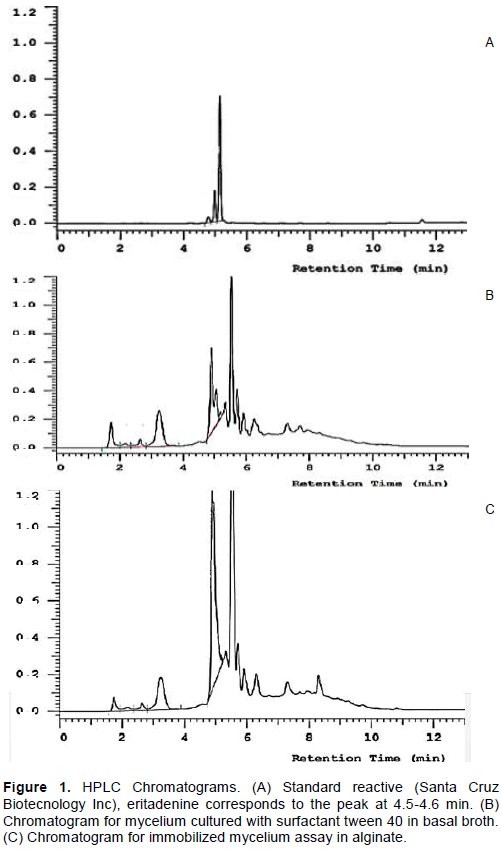
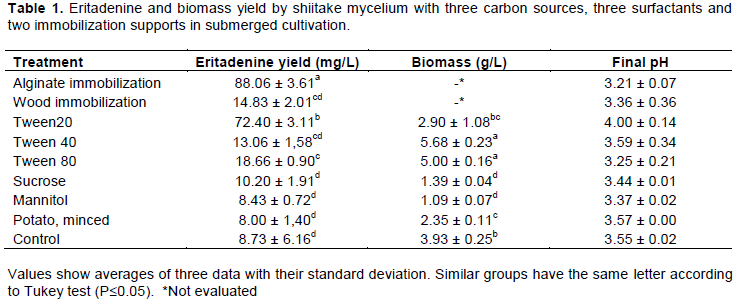
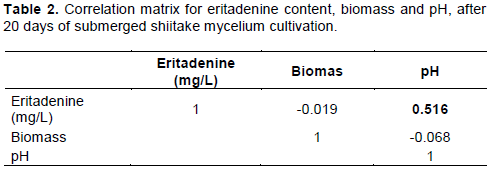
CONCLUSION
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
REFERENCES
|
Alaoui SM, Merzouki M, Penninckx MJ, Benlemlih M (2008). Relationship between cultivation mode of white rot fungi and their efficiency for olive oil mill wastewaters treatment. Electronic Journal of Biotechnology 11(4):1-8. |
|
|
Afrin S, Rakib MA, Kim BH, Kim JO, Ha YL (2016). Eritadenine from edible mushrooms inhibits activity of angiotensin converting enzyme in vitro. Journal of Agricultural Food Chemistry 64 (11):2263-8. |
|
|
Aminuddin H, Khan MA, Abidin H, Madzlan K, Suri R, Kamal MK (2007). Optimization of submerged culture for the production of Lentinula edodes mycelia biomass and amino acid composition by different temperatures. Journal of Tropical Agriculture and Food Science 35(1):131-138. |
|
|
Aminuddin HA, Khan M, Madzlan K (2013). Effects of pH on mycelial growth and amino acid composition of Lentinula edodes in submerged cultures. Journal of Tropical Agriculture and Food Science 41(1):63-70. |
|
|
Beltrán-García MJ, Orozco A, Samayoa I, Ogura T (2001). Lignin degradation products from corn stalks enhance notably the radial growth of basidiomycete mushroom mycelia. Revista de la Sociedad Química de Mexico 45:77-81. |
|
|
Bisen PS, Baghel RK, Sanodiya BS, Thakur GS, Prasad GBKS (2010). Lentinus edodes: A macrofungus with pharmacological activities. Current Medicinal Chemistry 17:2419-2430. |
|
|
ÄŒtrnáctá V, Fritzler JM, Šurinová M, Hrdý I, Zhu G, Stejskal F (2010). Efficacy of S-adenosylhomocysteine hydrolase inhibitors, D-eritadenine and (S)-DHPA, against the growth of Cryptosporidium parvum in vitro. Experimental Parasitology 126(2):113-116. |
|
|
Daza A, Manjón JL, Camacho M, Romero de la Osa L, Aguilar A, Santamaría C (2006). Effect of carbon and nitrogen sources, pH and temperature on in vitro culture of several isolates of Amanita caesarea (Scop.:Fr) Pers. Mycorrhiza 16(2):133-136. |
|
|
El-Refai HA, Shafei MS, Mostafa H, El-Refai AMH, Araby EM, El-Beih FM, Easa SM, Gomma SK (2016). Comparison of free and immobilized L-asparaginase synthesized by Gamma-Irradiated Penicillium cyclopium. Polish Journal of Microbiology 65(1):43-50. |
|
|
Enman J, Hodge D, Berglund KA, Rova U (2008). Production of the bioactive compound eritadenine by submerged cultivation of Shiitake (Lentinus edodes) mycelia. Journal of Agricultural and Food Chemistry 56:2609-2612. |
|
|
Enman J, Hodge D, Berglund KA, Rova U (2012). Growth promotive conditions for enhanced eritadenine production during submerged cultivation of Lentinus edodes. Journal of Chemical Technology and Biotechnology 87(7):903-907. |
|
|
Enman J, Patra A, Ramser K, Rova U, Berglund KA (2011). Solid state characterization of sodium eritadenate. American Journal of Analytical Chemistry 2:164-173. |
|
|
Gibbs PA, Seviour RJ, Schmid F (2000). Growth of filamentous fungi in submerged culture: Problems and possible solutions. Critical Reviews in Biotechnology 20(1):17-48. |
|
|
Gil-Ramirez A, Morales D, Soler-Rivas C (2018). Molecular actions of hypocholesterolemic compounds from edible mushrooms. Food and Function 9:53-69. |
|
|
Gonzales PL, Fari-a L, Carrau F, Batista-viera F, Brena BM (2014). Aroma enhancement in wines using co-immobilized Aspergillus niger glycosidases. Food Chemistry 143:185-191. |
|
|
Hadibarata T, Kristanti RA (2013). Effect of surfactants and identiï¬cation of metabolites on the biodegradation of flRA (2013). by basidiomycetes fungal isolate Armillaria sp. F022. Bioprocess and biosystems engineering 37(4):593-600. |
|
|
Hameed A, Hussain SA, Yang J, Umair MI, Liu Q, Ansar HRS, Song Y (2017). Antioxidants potential of the filamentous fungi (Mucor circinelloides). Nutrients 9(1101): 1-20. |
|
|
Hakala TK, Lundell T, Galkin S, Maijala P, Kalkkinen N, Hatakka A (2005). Manganese peroxidases, laccases and oxalic acid from the selective white rot fungus Physisporinus rivulosus grown on spruce wood chips. Enzyme and Microbial Technology 36(4):461-468. |
|
|
Hassegawa RH, Kasuya MCM, Vanetti MCD (2005). Growth and antibacterial activity of Lentinula edodes in liquid media supplemented with agricultural wastes. Electronic Journal of Biotechnology 8:212-217. |
|
|
Harris-Valle CM, Esqueda A, Sanchez M, Beltrán-García E, Valenzuela S (2007). Polar vineyard pruning extracts increase the activity of the main lignolytic enzymes in Lentinula edodes cultures. Canadian Journal of Microbiology 53:1150-1157. |
|
|
Hwang JA, Hossain ME, Yun DH, Moon ST, Kim GM, Yang CJ (2012). Effect of shiitake [Lentinula edodes (Berk.) Pegler] mushroom on laying performance, egg quality, fatty acid composition and cholesterol concentration of eggs in layer chickens. Journal of Medicinal Plants Research 6(1):146-153,159. |
|
|
Jakovljevic V, Milicevic J, Stojanovic J (2014). Detergent-like stressor and nutrient in metabolism of Penicillium chrysogenum. Biotechnology and Biotechnological Equipment 28(1):43-51. |
|
|
Jang Y, Park J, Ryoo R, Park Y, Ka KH (2016). Ergothioneine Contents of Shiitake (Lentinula edodes) Fruiting Bodies on Sawdust Media with Different Nitrogen Source. Korean Journal of Medical Mycology 44(2):100-102. |
|
|
Kasinath A, Novotny C, Svobodová K, Patel KC, Sasek V (2003). Decolorization of synthetic dyes by Irpex lacteus in liquid cultures and packed-bed bioreactor. Enzyme and Microbial Technology 32:167-173. |
|
|
Kirby J, Nishimoto M, Chow RWN, Pasumarthi VN, Chan R, Chan LJG, Petzold CJ, Keasling JD (2014). Use of nonionic surfactants for improvement of terpene production in Saccharomyces cerevisiae. Applied and Environmental Microbiology 80(21):6685-6693. |
|
|
Papagianni M, Joshi N, Moo-Young M (2002). Comparative studies on extracellular protease secretion and glucoamylase production by free and immobilized Aspergillus niger cultures. Journal of Industrial Microbiology and Biotechnology 29(5):259-263. |
|
|
Lazim ZM, Hadibarata T (2016). Lignolytic fungus Polyporus sp. S133 mediated metabolic degradation of fluorine. Brazilian Journal of Microbiology 47(3):610-616. |
|
|
Lopez-Pe-a D, Gutiérrez A, Esqueda M (2013). Cinética de crecimiento y composición química del micelio de Lentinula edodes cultivado en medio líquido suplementado con extractos de madera de vid. Revista Mexicana de Micología 37: 51-59. |
|
|
Loyola LN, Oyarce EC, Acu-a CC (2010). Evaluación del contenido de almidón en papas (Solanum tuberosum, sp. tuberosum cv. desirée), producidas en forma orgánica y convencional en la provincia de curicó, región del maule. IDESIA 28(2):41-52. |
|
|
Mahamud MA, Ohmasa M (2008). Effects of cultural conditions on high temperature tolerance of Lentinula edodes mycelia. Pakistan Journal of Biological Sciences 11(3):342-350. |
|
|
Mantle PG (2009). The role of tryptophan as a biosynthetic precursor of indole-diterpenoid fungal metabolites: Continuing a debate. Phytochemistry 70(1):7-10. |
|
|
Mwamburi LA, Laing MD, Miller RM (2015). Effect of surfactants and temperature on germination and vegetative growth of Beauveria bassiana. Brazilian Journal of Microbiology 46(1):67-74. |
|
|
Noreen S, Asgher M. Hussain F, Iqbal A (2016). Performance improvement of Ca-Alginate beads Cross-Linked Laccase from Trametes versicolor IBL-04. Bioresources 11(1):558- 572. |
|
|
O’sulivan SM, Woods JA, O’Brien NM (2004). Use of tween 40 and tween 80 to deliver a mixture of phytochemicals to human colonic adenocarcinoma cell (CaCo-2) monolayers. British Journal of Nutrition 91:757-764. |
|
|
Pedri ZC, Lozanob LMS, Hermannc KL, Helmd CV, Peraltae RM, Tavaresf LBB (2015). Influence of nitrogen sources on the enzymatic activity and grown by Lentinula edodes in biomass Eucalyptus benthamii. Brazilian Journal of Biology 75 (4):940-947. |
|
|
Quaicoe EH, Amoah C, Obodai M, Odamtten GT (2014). Nutrient requirements and environmental conditions for the cultivation of the medicinal mushroom (Lentinula Edodes) (Berk.) in Ghana. International Journal of Scientific and Technology Research 3(12):45-50. |
|
|
Rasmy GE, Botros WA, Kabeil SS, Daba AS (2010). Preparation of glucan from Lentinula edodes edible mushroom and elucidation of its medicinal value. Australian Journal of Basic and Applied Sciences 4(11):5717-5726. |
|
|
Rodriguez SC (2009). Dye removal by immobilized fungi. Biotechnology Advances 27:227-235. |
|
|
Sambamurthy K, Nageswarra L (1971). Improved medium for saprophytic production of ergot alkaloids by Claviceps purpurea (Fr.) Tul. Biotechnology and Bioengineering 13: 331-334. |
|
|
Shide EG, Wuyep PA, Nok AJ (2004). Studies on the degradation of wood sawdust by Lentinus squarrosulus (Mont.) Singer. African Journal of Biotechnology 3(8):395-398. |
|
|
Shu-Lei W, Jing-Yu L, Qing-Jiu T (2012). Advances in studies of eritadenine. Mycosistema 31(2):151-158. |
|
|
Shimada Y, Morita T, Sugiyama K (2003). Eritadenine-induced alterations of plasma lipoprotein lipid concentrations and phosphatidylcholine molecular species profile in rats fed cholesterol free and cholesterol enriched diets. Bioscience, biotechnology, and biochemistry 67(5):996-1006. |
|
|
Peksen A, Kibar B, Yakupoglu G (2013). Favourable culture conditions for mycelial growth of Hydnum repandum, a medicinal mushroom. African Journal of Traditional, Complementary and Alternative Medicines 10 (6):431-434. |
|
|
Stamets P (2000). Growing gourmet and medicinal mushrooms, USA. Ten Speed Press. CA. |
|
|
Susla M, Novotny C, Svobodová K (2007). The implication of Dichomitus squalens laccase isoenzymes in dye decolorization by immobilized fungal cultures. Bioresource Technology 98:2109-2115. |
|
|
Tepwong P, Giri A, Ohshima T (2012a). Effect of mycelial morphology on ergothioneine production during liquid fermentation of Lentinula edodes. Mycoscience 53:102-112. |
|
|
Tepwong P, Giri A, Sasaki F, Fukui R, Ohshima T (2012b). Mycobial enhancement of ergothioneine by submerged cultivation of edible mushroom mycelia and its application as an anti-oxidative compound. Food Chemistry 131:247-258. |
|
|
Ünal A, Kolankaya N (2013). Determination of optimum immobilization conditions of Trametes versicolor laccase with sodium alginate beads. IUFS Journal of Biology 72(2):15-21. |
|
|
Weber SS, Polli F, Boer R, Bovenberg RAL, Driessena AJM (2012). Increased penicillin production in Penicillium chrysogenum production strains via balanced overexpression of isopenicillin N acyltransferase. Applied and Environmental Microbiology 78(19):7107-7113. |
|
|
Yang H, Hwang I, Kim S, Ahn C, Hong EJ, Jeung EB (2013). Preventive effects of Lentinus edodes on homocysteinemia in mice. Experimental and Therapeutic Medicine 6:465-468. |
|
|
Yamada T, Komoto J, Lou K, Ueki A, Hua DH, Sugiyama K, Takata Y, Ogawa H, Takusagawa F (2007). Structure and function of eritadenine and its 3-deaza analogues: potent inhibitors of S-adenosylhomocysteine hydrolase and hypocholesterolemic agents. Biochemical Pharmacology 173(7):981-989. |
|
|
Zagrean V, Neata G, Stanciulescu B (2017). Influence of temperature on mycelial growth of some Pleurotus eryngii and Lentinula edodes strains in vitro. Bulletin UASVM Horticulture 74(1):81-82. |
|
|
Zhang YP, Jiao RH, Lu YH, Yao LY (2016). Improvement of chaetominine production by tryptophan feeding and medium optimization in submerged fermentation of Aspergillus fumigatus CY018. Bioresource Bioprocess 3:45. |
|
|
Zhang J, Wang YL, Lu LP, Zhang BB, Xu GR (2014). Enhanced production of monacolin K by addition of precursors and surfactants in submerged fermentation of Monascus purpureus 9901. Biotechnology and Applied Biochemistry 61(2):202-207. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


