ABSTRACT
In this study lactic acid bacteria were investigated for their presence in fresh palm wine and their effects on humoral immunity examined. The bacteria were isolated from fresh palm wine in a tenfold serial dilution. The isolates were purified by repeated subculture on De Man Rogosa Sharpe (MRS) agar and characterized phenotypically and genotypically. The identified Lactobacillus spp. were investigated for their effects on antibody (humoral immunity) - IgG, IgG1 and IgG2a secretion using sheep red blood cell as antigen (SRBC). Lactobacillus spp. were identified as Lactobacillus brevis, Lactobacillus paracasei subsp. tolerans, Lactobacillus paracasei and Lactobacillus yonginensis. The isolates produced no significant effect on IgG antibody after 4 days’ post-secondary challenge. However, they had significant percentage stimulation at 9 days post-secondary challenge of 100.2, 122.9, 106.4 and 118.3% for L. brevis, L. paracasei sub. tolerans, L. paracasei, and L. yonginensis, respectively. The isolates had marginal and somewhat suppressive effects on IgG1 and IgG2a at both 4 and 9 days’ post-secondary challenge. The results show that fresh palm wine contains Lactobacillus spp. capable of stimulating antibodies production.
Key words: Humoral immunity, antibody, palm wine, Lactobacillus spp.
Lactobacilus spp. are important class of bacteria that have been extensively studied by scientists in recent times. They are generally regarded as safe (Gras) bacteria hence exploited in several ways to the benefit of man. Lactobacillus have been linked with the production of antimicrobial substances, antibiotic resistance patterns, improving digestive ability and antibody mediated response with demonstrable efficacy and safety (Hou et al., 2015; Wang et al., 2012). Studies over the years have proved that Lactobacillus spp are potent modulators of the immune system.
They are able to modulate both the adaptive and innate components of the immune system through the regulation of the functions of dendritic cells, macrophages, T and B lymphocytes. The success of this class of bacteria as an immunomodulator is linked to its ability to bind pattern recognition receptors (PRR) expressed on immune cells. These PRR in turn recognize conserved molecular structure known as microbe associated molecular patterns (MAMPS) and signal to induce the production of cytokines, chemokines and other innate effectors (Wells et al., 2010; Abreu, 2010; Kawai and Akira, 2010). Recent studies have shown that probiotics (e.g lactic acid bacteria) are becoming popular as an option in the treatment of inflammatory disorders (Evrad et al., 2011; Kwon et al., 2010). They also influence positively though indirectly, the regulatory T cells (Treg) by providing a favourable environment (Petersen et al., 2012). Lactobacillus spp administration/studies in vivo and ex vivo produced significant increase in all functions of peritoneal macrophages viz, increased microbiocidal and phagocytic activities and increased production of cytokines induced by macrophages (Marranzino et al., 2012). Lactobacillus lactis strains commonly found in food products directly stimulate plasmacytoid dendritic cells (pDC) resident in the intestinal draining mesenteric lymph nodes to produce not only type 1 IFN but also IFN-λ and augment the capacity of pDC to induce CD4+CD25+Treg generation (Jounai et al., 2012). Lactobacillus acidophilus increases the cytotoxic activity of natural killer cells (Corthesy et al., 2007). These beneficial and safe bacteria stimulate innate immune system in immune deficient individuals (Delcenserie et al., 2008) and also in immune deficient elderly persons where they stimulate both phagocytic activity of macrophages and natural killer cell function (El-Gaaly et al., 2016).
It is an established fact that Lactobacillus spp belong to a group of bacteria that have been extensively studied because of their enormous health benefits. The lyophilized forms of these bacteria are now used as therapeutics in the management of diarrhea. This study derived it uniqueness from the fact that these bacteria was isolated from a natural alcoholic beverage. Elaeis guineensis (Palm tree) sap (Palm wine) was studied for the presence of these important bacteria and their effect on humoral immunity. Palm wine is a whitish liquid produced as a result of natural fermentation by acetic acid bacteria, lactic acid bacteria and yeast (Santiago-Urbina and Ruiz-Teran, 2014). These organisms usually cause the breakdown of sugar in the sap into alcohol and other products (Obire, 2005). These organisms are responsible for the sour taste experienced on this beverage over time as the tapping process progresses. The chemistry of what happens is that at the point of tapping the beverage is rich in sugar hence very sweet. The activities of these beneficial bacteria tend to convert the sweet beverage into alcohol and ethanoic acid (sour taste experienced after about 24 h of fermentation) in a time dependent process (Eze et al., 2019). In West African sub-regions, this sweet and fresh alcoholic beverage is linked with some health benefits such as increased sperm in men and breast milk production in women, and high content of B1, B2, B3, and B6 vitamins (Mbuagbaw and Noorduyn, 2012). The beverages also possess high content of amino acid, potassium, zinc and iron (Carousel, 2015).
In this study, we investigated the presence of Lactobacillus as they are suspected to be involved in the fermentation of palm wine and the effect of the isolates on humoral immunity was determined.
Sources of Lactobacillus
Fresh palm wine from oil palm tree Elaeis guineensis, Linex capsule (lyophilized lactobacillus capsule) Sandoz Pharmaceutical Slovenia.
Sources of antibodies and antigens used
Ovalbumin (Sigma-Aldrich, USA), Goat anti-mouse IgGFab HRP (Southern biotech, USA), Goat anti-mouse IgG1Fab HRP (Southern biotech, USA), Goat anti-mouse IgG2aFab HRP (Southern biotech, USA), albino mice (28-30 g), Sheep red blood cell (SRBC).
Collection of palm wine
The fresh palm wine samples were collected at about 6:30 am by Mr Anthony Idoko a local Palm wine tapper from Onicha Enugu Ezike, Igbo- Eze North Local government area of Enugu State, Nigeria. The samples were kept under cold conditions using ice pack to reduce the rate of fermentation while being transported to the laboratory.
Preparation of media and isolation of Lactobacillus spp
The media were prepared following the manufacturer`s specifications. The test organisms were isolated from Palm wine using quadrant streak plate method. A wire loop was used to collect a loopfull of the homogenized ten- fold serial dilution of the fresh Palm wine samples and streaked on the surface of sterile modified MRS media under aseptic conditions. The inoculated media were incubated using anaerobiosis generator at 37°C for 24-48 h. After the incubation, distinct colonies were subcultured on MRS agar to obtain pure cultures. The purified isolates were streaked on MRS agar slants and stored at 4°C for further use.
Phenotypic characterization of the culture
The isolates were presumptively identified based on cultural, morphological and some biochemical characteristics. The parameters investigated included colony morphology, Gram reactions, endospore formation, catalase production, motility, and sugar fermentation. The results were compared to Holt et al. (1994) Bergey's Manual of Determinative Bacteriology.
Genotypic characterization of the Isolates
DNA extraction and polymerase chain reaction (PCR) amplification
DNA extraction was carried out on test organisms’ isolated from palm wine using the Jena Bioscience Bacteria DNA Preparation Kit (http://www.jenabioscience.com). Polymerase chain reaction was carried out to identify the suspected lactic acid bacteria isolated from palm wine using the primer pair BSF8 (AGAGTTTGATCCTGGCTCAG) and BSR534 (ATTACCGCGGCTGCTGC). The primer pair are lactic acid specific. The PCR reaction was carried out using the Solis Biodyne 5X HOT FIREPol Blend Master mix. PCR was performed in 25 µl of a reaction mixture, and the reaction concentration was brought down from 5x concentration to 1X concentration containing 1X Blend Master mix buffer Buffer (Solis Biodyne), 1.5 mM MgCl2, 200 µM of each deoxynucleoside triphosphates (dNTP)(Solis Biodyne), 25 pMol of each primer (BIOMERS, Germany), 2 unit of Hot FIREPol DNA polymerase (Solis Biodyne), Proofreading Enzyme, 5 µl of the extracted DNA, and sterile distilled water was used to make up the reaction mixture. Thermal cycling was conducted in a Peltier thermal cycler (PTC100) (MJ Research Series) for an initial denaturation of 95°C for 15 min, followed by 35 amplification cycles of 30 s at 95°C; 1 min. at 58°C and 1 min 30 s at 72°C. This was followed by a final extension step of 10 min at 72°C. The amplification product was separated on a 1.5% agarose gel and electrophoresis was carried out at 80V for 1 h 30 min. After electrophoresis, DNA bands were visualized using ethidium bromide staining. 100 bp DNA ladder was used as DNA molecular weight standard.
Identification of the isolates
All the isolates were identified using 16S rRNA. All PCR products were purified and sent to Epoch Life science (USA) for Sanger sequencing. The corresponding sequences were identified using the online blast search at
http://blast.ncbi.nlm.nih.gov/Blast.cgi.
Immunological studies
Grouping and dosing of the animals
The animals were grouped into 6 groups of 5 animals (mice) per group. The first 4 groups received 0.2 ml of 3×107 cfu/ml of each of the isolates determined by 0.5 McFarland standard. The 5th and the 6th group represent positive and negative control groups respectively. The positive control received linex capsule (0.2 ml) a brand of lyophilized Lactobacillus spp. at a dose of 1.2 × 105 cfu/ml.
Studies on humoral antibody HA response
This is a slight modification of the method by Sharma et al. (1996). Mice were immunized by an intraperitoneal (ip) injection of 0.2 ml of 1×109 SRBC ml-1 on (day 0) and challenged by similar i.p injection of the same amount on day 5. The animals were bled and the serum analysed for the antibody titre by ELISA on days 10 and 15. Briefly 1 mg/ml of ova in bicarbonate buffer (pH 9.6) was prepared and used to coat the 96 well ELIZA plates and the plates incubated at 4°C for about 14 h. The plates were washed with PBS-Tween (0.05% Tween-20 in 0.1 M PBS). The plates were blocked with 20 μl/well of 5% v/v solution of fat free milk in PBS-Tween and incubated at room temperature for 1 h. The plates were washed 3 times with PBS-Tween. The sera from the animals were diluted 1:20 with 2% fat free milk and added 100 μl/well in duplicates. The plates were washed with PBS-Tween. 100 μl of 1:1000 dilution of HRP-antigoat Immunoglobulin were added to the wells and incubated for 1 h at room temperature. The plates were washed four times with PBS-Tween. 100 μl of the quenching agent TMT were added per well and allowed for at least 15-20 min and the plates read in ELIZA machine at 405 nm. The LAB isolates were administered 3 days prior to immunization and continued on a daily dose of 3×107 cfu/ml until the 14th day. The immunoglobulin studied included IgG, IgG1 and IgG2a.
Statistical analysis
The statistical analysis was done using Graph Pad prism version 5.0. One-way ANOVA followed by Post-hoc Dunnet were used to compare mean ± SEM, and values were considered significant at p < 0.05.
The morphological and biochemical characteristics of the isolated Lactobacillus
The morphological and biochemical characteristics of the isolated Lactobacillus are as shown in Table 1.
Genotypic and blasted sequence results of the isolates
The result of the blasted sequenced showed the presence of four species of Lactobacillus namely: L. brevis, L. paracasei subsp. Tolerans, L. paracasei and L. yonginensi. Primer pairs - BSF8 (AGAGTTTGATCCTGGCTCAG) and BSR534 (ATTACCGCGGCTGCTGC) was used to identify the suspected lactic acid bacteria isolated from the palm wine.
The effect of Lactobacillus spp. on antibody secretion
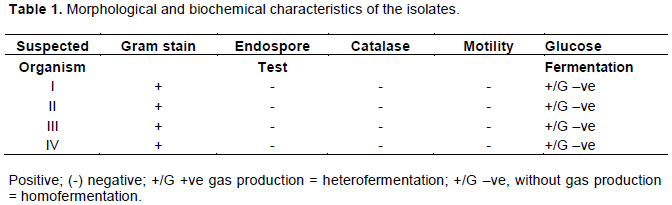
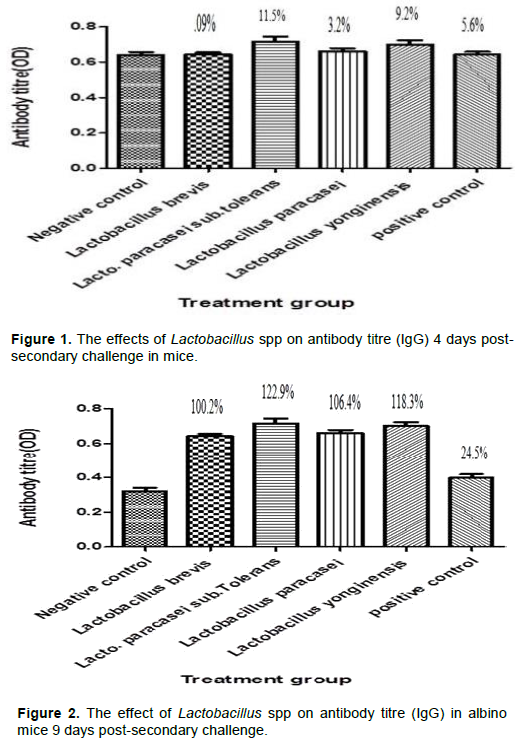
Figures 1 and 2 shows the effect of Lactobacillus spp on IgG antibody secretion 4 days and 9 days post-secondary challenge in mice. The isolates produced insignificant effects on IgG antibody secretion 4 days post-secondary challenge; however, there was a significant increase of 100.2, 122.9, 106.4 and 118.3% for L. brevis, L. paracasei subsp. Tolerans, L. paracasei and L. yonginensi respectively when compared with the negative control. The effects of Lactobacillus spp on IgG1 and IgG2a secretion 4 and 9 days post-secondary challenge is as shown in Figures 3 to 6.
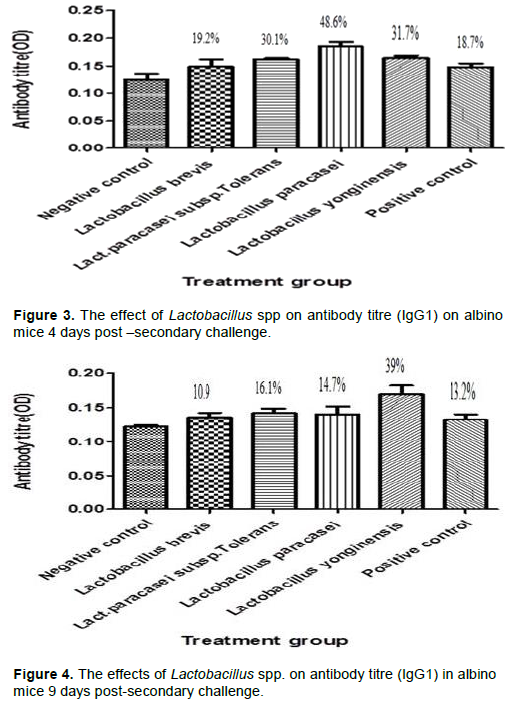
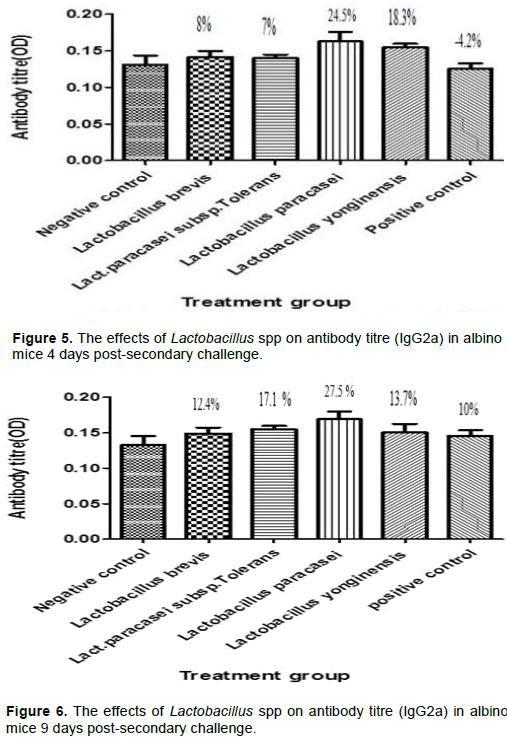
L. paracasei subsp.Tolerans, L. paracasei and L. yonginensis produced a significant percentage increase of 30.1, 48.6 and 31.7% on the albino mice 4 days post-secondary challenge while only L. yonginensis produced a significant percentage increase of 39% 9 days post-secondary challenge when compared with the negative control. In the case of IgG2a there was no significant percentage stimulation at 4 and only L. paracasei produced significant percentage stimulation at 9 days post challenge.
Humoral immunity refers to antibody-mediated immune responses. Antibodies are produced by plasma cells and protect the host from infection in three main ways: by binding to pathogens to inhibit their toxic effects or infectivity (neutralization), by coating pathogens and facilitating their uptake and killing by phagocytes (opsonization) and by activating the complement cascade. Immunoglobulin G (Ig) is basically a major component of the circulating immunoglobulin. In mice it is sub classified into IgG1, IgG2a, IgG2b and IgG3 (Pelsue, 2019). The genotypic characterization and blasted sequence result confirmed the presence of Lb. brevis, Lb. paracasei sub. Tolerans, Lb. paracasei and Lb. yonginensis. The effects of the different Lactobacillus isolates on IgG, IgG1 and IgG2a secretion were investigated at 4 and 9 days post- secondary challenge on albino mice. The serum was analyzed for the antibody titre post-secondary challenge because IgM is the predominant immunoglobulin after initial exposure or post primary challenge. The result of the ELISA showed that there was an insignificant increase in the antibody titre for the different Lactobacillus species 4 days post-secondary antigenic challenge for IgG (Figure 1). However at 9 days post-secondary antigenic challenge for IgG, there was marked increase of 100.2, 122.9, 106.4 and 118.3% for Lb. brevis, Lb. paracasei sub. Tolerans, Lb. paracasei, and Lb. yonginensis, respectively (Figure 2). The non-significant effect experienced 4 days post challenge could be explained from the fact that the antibody secretion triggered by Lactobacillus may be T cell dependent. Antibody production that is T cell dependent involves a cascade of events thus: activated B and T cells remain complexed for 3 days and an exchange of signals, such as CD40/CD40L, leads to initiation of one of two fates for the B cell, some naïve B cells move into the extra follicular region of the lymph node, where they differentiate into short-lived Plasma cells that produce only IgM as an initial response to infection while others migrate into the follicles where they form germinal centers (GC) (Paus et al., 2006). The GC is the site where B cells undergo affinity maturation, clonal expansion, and ultimately differentiate into high affinity, long lived plasma cells or memory B cells (Jackson and Elsawa, 2015). The sera of the experimental mice analyzed at 4 days post-secondary challenge showed that Lb. brevis produced an insignificant effect on IgG1 secretion while Lb. paracasei sub. Tolerans, Lb. paracasei, and Lb. yonginensis produced significant percentage increase as shown in Figure 3. The effect of the isolates on IgG1 9 day’s post-secondary challenge showed that all the three isolates had insignificant effects on antibody production except Lb. yonginensis (Figure 4). Figures 5 and 6 showed that all the isolates produced no significant effect on IgG2a 4 days post- secondary challenge, however only L. paracasei produced significant antibody titre 9 days post challenge while the other three produced no significant titre.
The result of this experiment is in agreement with previous studies on the ability of Lactobacillus spp to stimulate antibody production (Davras et al., 2018; Easo et al., 2002). A comparison of the effects of the different isolates on the studied immunoglobulins, showed that the isolates produced better percentage stimulation of IgG and IgG1 when compared with the overall effect on IgG2a. This could be because in mice, IgG1 and IgE have been widely used as the surrogate markers of humoral antibody (T-helper [Th2] activation) responses as IL-4 secreted by Th2 cells induces Ig class switching into IgG1 and IgE subclasses (Wang et al., 2007). IL-4 plays an important role in antibody production by inducing the proliferation and differentiation of B cells into plasma cells. On the other hand, IgG2a and IgG3 is the surrogate markers of cellular immune response (Th1 activation) as IFN-γ produced by Th1 cells induces Ig class switching into IgG2a or IgG3 subclasses (Stevens et al., 1988).
Palm wine has shown that it is an alcoholic beverage with a difference since it is a natural habitat for immune enhancing, beneficial and generally regarded as safe (GRAS) bacteria, Lactobacillus. The four Lactobacillus spp were isolated from the fresh and sweet (high sugar content) beverage immediately (0-4 h) after fermentation. Therefore, since fresh palm wine has in abundance these beneficial bacteria, the locals who indulge in excessive consumption of fermented and soured palm wine should be educated on the health benefits of the fresh drink. This will enable them avoid the deleterious effects of alcohol while benefiting maximally from the enormous health benefit of this natural beverage.
Fresh E. guinneesis sap (palm wine) is home to Lactobacillus spp that has the capacity to positively stimulate the production of antibodies in mice.
The authors are proposing an increased research on this natural beverage especially as it concerns delay/control of fermentation by these beneficial bacteria. This will automatically transform this beverage to a healthier drink than alcoholic.
The authors have not declared any conflict of interests.
REFERENCES
|
Abreu MT (2010). Toll-like receptor signalling in the intestinal epithelium: how bacterial recognition shapes intestinal function. Nature Review Immunology 10(2):131-144.
Crossref
|
|
|
|
Carousel JP (2015). Information-and-health-benefits-of-palm-wine. Retrieved September 17, 2017, from
View
|
|
|
|
|
Corthesy B, Rex Gaskins H, Mercenier A (2007). Cross-Talk between probiotic bacteria and the host immune system. American Society for Nutrition 137:781S-790S.
Crossref
|
|
|
|
|
Davras F, Guzel-Seydian ZB, Kok Tas T (2018). Immunological effects of Kefir produced from Kefir grains versus starter cultures when fed to mice. Functional Foods in Health and Diseases 8(8):412-423.
Crossref
|
|
|
|
|
Delcenserie V, Martel D, Lamoureux, Amiot J, Boutin Y, Roy D (2008). Immunomodulatory effects of probiotics in the intestinal tract. Current Issues in Molecular Biology 10:37-54.
|
|
|
|
|
Easo JG, Measham JD, Munroe J, Green-Johnson JM (2002). Immunostimulatory actions of lactobacilli: Mitogenic induction of antibody production and spleen cell proliferation by lactobacillus delbrueckii subsp. bulgaricus and lactobacillus acidophilus. Food and Agricultural Immunology 14:73-83.
Crossref
|
|
|
|
|
EL-Gaaly SA, Radwan HH, Attiha MY (2016). Immunomodulatory factor of Lactobacillus acidophilus in pathogenesis of chronic HCV. The Egyptian Journal of Hospital Medicines 63:229-237.
Crossref
|
|
|
|
|
Evrad B, Coudeyras S, Dosgilbert A, Charbonnel N, Alame J, Tridon A, Forestier C (2011). Dose-dependent immunomodulation of human dendritic cells by the probiotic lactobacillus rhamnosus LCr 35. PLoS ONE 6(4):e18735.
Crossref
|
|
|
|
|
Eze CO, Berebon DP, Gugu TH, Nworu CS, Esimone CO (2019). Effects of lactobacillus spp isolated from the sap of palm tree Elaeis guineensis (palm wine) on cellular and innate immunity. African Journal of Microbiology Research 13(2):33-39.
Crossref
|
|
|
|
|
Holt JG, Kreig NR, Sneath PHA, Stanely JT, Williams ST (1994). Bergeys Manual of Determinative Bacteriology-Ninth Edition. Lippincott, Williams and Wilkins, Baltimore 787p.
|
|
|
|
|
Hou C, Zeng X, Yang F, Liu H, Qiao S (2015). Study and the use of the probiotic Lactobacillus reuteri in pigs: A review. Journal of Animal Science and Biotechnology 6:14.
Crossref
|
|
|
|
|
Jackson DA, Elsawa SF (2015). Factors regulating immunoglobulin production by normal and disease associated plasma cells. Biomolecules 5(1):20-40.
Crossref
|
|
|
|
|
Jounai KI, Ikado K, Sugimura T, Ano Y, Braun J, Fujiwara D (2012). Spherical lactic acid bacteria activate plasmacytoid dendritic cells immunomodulatory function via The TLR9-dependent crosstalk with myeloid dendritic cell. PLoS ONE 7(4):e32588.
Crossref
|
|
|
|
|
Kawai T, Akira S (2010). The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nature Immunology 11(5):373-384.
Crossref
|
|
|
|
|
Kwon HC, Lee CG, So JS, Chae CS, Hwang JS, Sahoo A, Nam JH, Rhee JH, Hwang KC, Im SH (2010). Generation of regulatory dendritic cells and CD4+ Foxp3+ T cells by probiotics administration suppresses immune disorders. Proceedings of the National Academy of Science of the USA 107:2159-2164.
Crossref
|
|
|
|
|
Marranzino G, Villena J, Salva S, Alvarez S (2012). Stimulation of macrophages by Immunobiotic Lactobacillus strains: Influence beyond the intestinal tract. Microbiology and Immunology 56:771-781.
Crossref
|
|
|
|
|
Mbuagbaw L, Noorduyn SG (2012). The palm wine trade: occupational and health hazards. The International Journal of Occupational and Environmental Medicine 3:157-164.
|
|
|
|
|
Obire O (2005). Activity of yeast species in palm sap obtained from three areas in Edo State, Nigeria. Journal of Applied Science Environmental Management 9:25-30.
|
|
|
|
|
Paus D, Phan TG, Chan TD, Gardam S, Basten A, Brink R (2006). Antigen recognition strength regulates the choice between extrafollicular plasma cell and germinal center B cell differentiation. Journal of Experimental Medicine 203:1081-1091.
Crossref
|
|
|
|
|
Pelsue S (2019). IgG function.
|
|
|
|
|
Petersen ER, Claesson MH, Schmidt EG, Jensen SS, Ravn P, Olsen J, Ouwehand AC, Kristensen NN (2012). Consumption of probiotics increases the effect of regulatory T cells in transfer colitis. Inflamatory Bowel Diseases 18(1):131-142.
Crossref
|
|
|
|
|
Santiago-Urbina JA, Ruiz-Teran F (2014). Microbiology and biochemistry of traditional palm wine produced around the world. International Food Research Journal 21(4):1261-1269.
|
|
|
|
|
Sharma ML, Singh B, Chandan BK, Khajuria A, Kaul A, Bani S, Banerjee SK, Gabbhur SS (1996). Action of some flavonoids on specific and non-specific immune mechanisms. Phytomedicine 3:191-195.
Crossref
|
|
|
|
|
Stevens TL, Bossie A, Sanders VM, Fernandez-Botran R, Coffman RL, Mosmann TR, Vitetta ES (1988). Regulation of antibody isotype secretion by subsets of antigen-specific helper T cells. Nature 334(6179):255-258.
Crossref
|
|
|
|
|
Wang J, Ji HF, Wang SX, Zhang DY, Liu H, Shan DC, Wang YM (2012). Lactobacillus plantarum ZLP 001: in vitro assessment of antioxidant capacity and effect on growth performance and antioxidant status in weaning piglets. Asian-Australasian Journal of Animal Science 25(8):1153-1158.
Crossref
|
|
|
|
|
Wang G, Zhao J, Liu J, Huang Y, Zhong JJ, Tang W (2007). Enhancement of IL-2 and IFN-gamma expression and NK cells activity involved in the anti-tumor effects of ganoderic acid Me in vivo. International Immunopharmacology 7(6):864-70.
Crossref
|
|
|
|
|
Wells JM, Loonen LM, Karczewski JM (2010). The role of innate signaling in the homeostasis of tolerance and immunity in the intestine. International Journal of Medical Microbiology 300(1):41-48.
Crossref
|
|